

On the independent origins of complex brains and neurons
- PMID: 20029182
- PMCID: PMC2855278
- DOI: 10.1159/000258665
On the independent origins of complex brains and neurons
Abstract
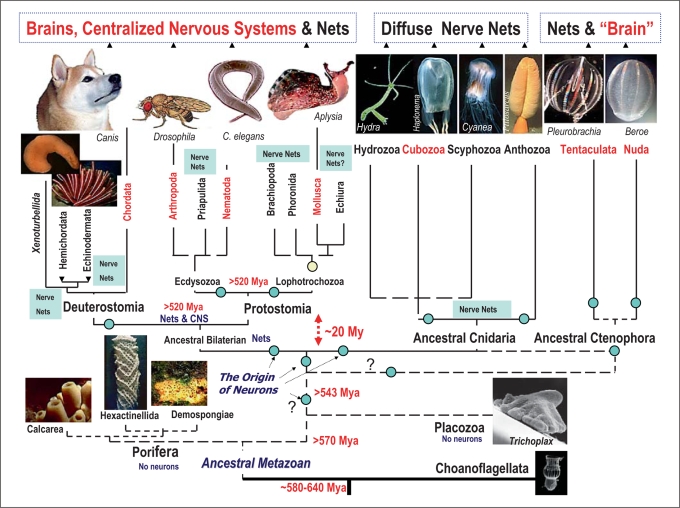
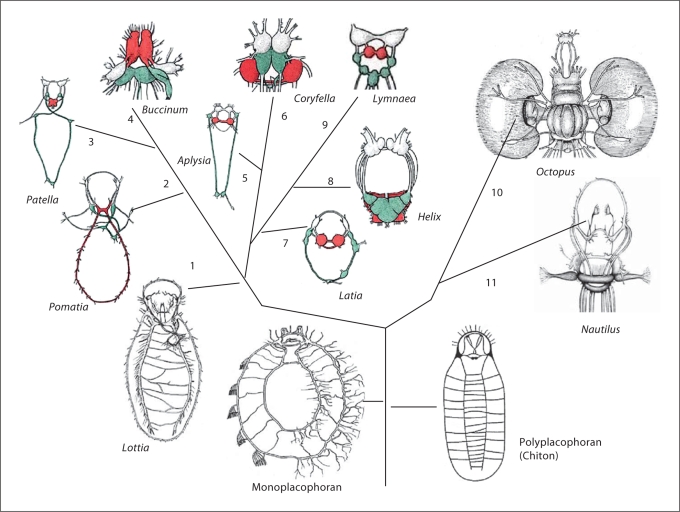
Analysis of the origin and evolution of neurons is crucial for revealing principles of organization of neural circuits with unexpected implications for genomic sciences, biomedical applications and regenerative medicine. This article presents an overview of some controversial ideas about the origin and evolution of neurons and nervous systems, focusing on the independent origin of complex brains and possible independent origins of neurons. First, earlier hypotheses related to the origin of neurons are summarized. Second, the diversity of nervous systems and convergent evolution of complex brains in relation to current views about animal phylogeny is discussed. Third, the lineages of molluscs and basal metazoans are used as illustrated examples of multiple origins of complex brains and neurons. Finally, a hypothesis about the independent origin of complex brains, centralized nervous systems and neurons is outlined. Injury-associated mechanisms leading to secretion of signal peptides (and related molecules) can be considered as evolutionary predecessors of inter-neuronal signaling and the major factors in the appearance of neurons in the first place.
Copyright 2009 S. Karger AG, Basel.
Figures
Comment in
-
Cladistic analysis reveals brainless urbilateria.Brain Behav Evol. 2010;76(1):1-2. doi: 10.1159/000316443. Epub 2010 Sep 30. Brain Behav Evol. 2010. PMID: 20926852 No abstract available.
References
-
- Arenas-Mena C, Cameron AR, Davidson EH. Spatial expression of Hox cluster genes in the ontogeny of a sea urchin. Development. 2000;127:4631–4643. - PubMed
-
- Arendt D. Genes and homology in nervous system evolution: comparing gene functions, expression patterns, and cell type molecular fingerprints. Theor Biosci. 2005;124:185–197. - PubMed
-
- Arendt D. The evolution of cell types in animals: emerging principles from molecular studies. Nat Rev Genet. 2008;9:868–882. - PubMed
-
- Arendt D, Nubler-Jung K. Inversion of dorsoventral axis? Nature. 1994;371:26. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
