

Comparative neurobiology of feeding in the opisthobranch sea slug, Aplysia, and the pulmonate snail, Helisoma: evolutionary considerations
- PMID: 20029185
- PMCID: PMC2855281
- DOI: 10.1159/000258668
Comparative neurobiology of feeding in the opisthobranch sea slug, Aplysia, and the pulmonate snail, Helisoma: evolutionary considerations
Abstract
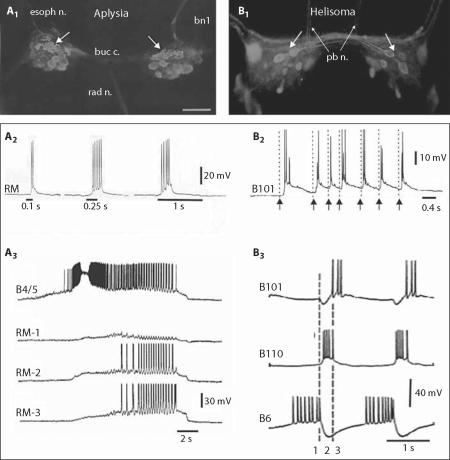
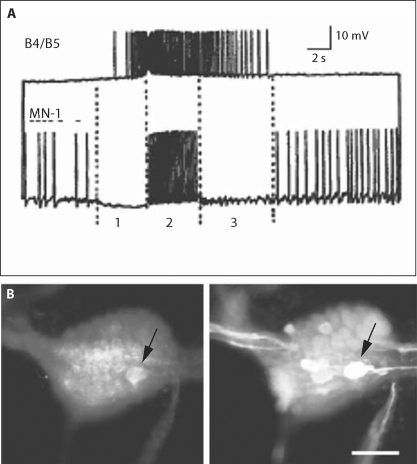
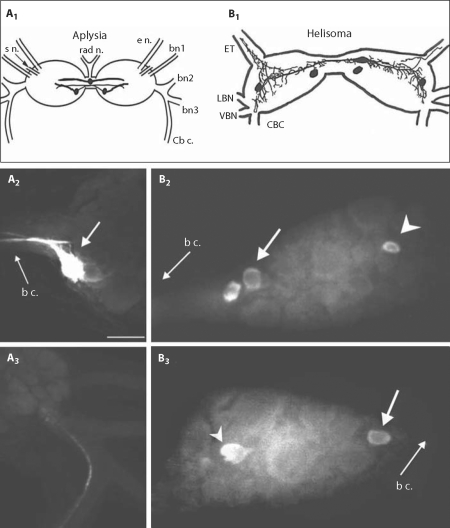
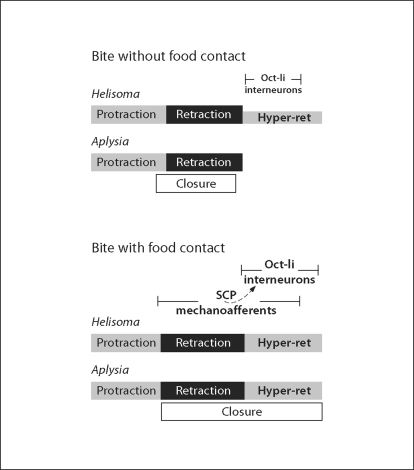
The motor systems that generate feeding-related behaviors of gastropod mollusks provide exceptional opportunities for increasing our understanding of neural homologies and the evolution of neural networks. This report examines the neural control of feeding in Helisoma trivolvis, a pulmonate snail that ingests food by rasping or scraping material from the substrate, and Aplysia californica, an opisthobranch sea slug that feeds by using a grasping or seizing motion. Two classes of neurons that are present in the buccal ganglia of both species are considered: (1) clusters of peptidergic mechanoafferent cells that transmit sensory information from the tongue-like radula/odontophore complex to the central motor circuit; and (2) sets of octopamine-immunoreactive interneurons that are intrinsic to the feeding network. We review evidence that suggests homology of these cell types and propose that their roles have been largely conserved in the control of food-scraping and food-grasping consummatory behaviors. We also consider significant differences in the feeding systems of Aplysia and Helisoma that are associated with the existence of radular closure in Aplysia, an action that does not occur in Helisoma. It is hypothesized that a major adaptation in the innervation patterns of analogous, possibly homologous muscles could distinguish the food-scraping versus food-grasping species. It appears that although core CPG elements have been largely conserved in this system, the neuromuscular elements that they regulate have been more evolutionarily labile.
Copyright 2009 S. Karger AG, Basel.
Figures

Similar articles
-
Diverse synaptic connections between peptidergic radula mechanoafferent neurons and neurons in the feeding system of Aplysia.J Neurophysiol. 2000 Mar;83(3):1605-20. doi: 10.1152/jn.2000.83.3.1605. J Neurophysiol. 2000. PMID: 10712483
-
Outputs of radula mechanoafferent neurons in Aplysia are modulated by motor neurons, interneurons, and sensory neurons.J Neurophysiol. 2000 Mar;83(3):1621-36. doi: 10.1152/jn.2000.83.3.1621. J Neurophysiol. 2000. PMID: 10712484
-
GABA-like immunoreactivity in Biomphalaria: Colocalization with tyrosine hydroxylase-like immunoreactivity in the feeding motor systems of panpulmonate snails.J Comp Neurol. 2018 Aug 1;526(11):1790-1805. doi: 10.1002/cne.24448. Epub 2018 May 6. J Comp Neurol. 2018. PMID: 29633264 Free PMC article.
-
Premotor neurons in the feeding system of Aplysia californica.J Neurobiol. 1989 Jul;20(5):497-512. doi: 10.1002/neu.480200516. J Neurobiol. 1989. PMID: 2664083 Review.
-
The Sea Slug, Pleurobranchaea californica: A Signpost Species in the Evolution of Complex Nervous Systems and Behavior.Integr Comp Biol. 2015 Dec;55(6):1058-69. doi: 10.1093/icb/icv081. Epub 2015 Jul 10. Integr Comp Biol. 2015. PMID: 26163678 Free PMC article. Review.
Cited by
-
Localization of tyrosine hydroxylase-like immunoreactivity in the nervous systems of Biomphalaria glabrata and Biomphalaria alexandrina, intermediate hosts for schistosomiasis.J Comp Neurol. 2014 Aug 1;522(11):2532-52. doi: 10.1002/cne.23548. Epub 2014 Apr 4. J Comp Neurol. 2014. PMID: 24477836 Free PMC article.
-
Octopamine promotes rhythmicity but not synchrony in a bilateral pair of bursting motor neurons in the feeding circuit of Aplysia.J Exp Biol. 2010 Apr;213(Pt 7):1182-94. doi: 10.1242/jeb.040378. J Exp Biol. 2010. PMID: 20228355 Free PMC article.
-
Neurotransmission and neuromodulation systems in the learning and memory network of Octopus vulgaris.J Morphol. 2022 May;283(5):557-584. doi: 10.1002/jmor.21459. Epub 2022 Feb 8. J Morphol. 2022. PMID: 35107842 Free PMC article.
-
GABA as a Neurotransmitter in Gastropod Molluscs.Biol Bull. 2019 Apr;236(2):144-156. doi: 10.1086/701377. Epub 2019 Jan 16. Biol Bull. 2019. PMID: 30933636 Free PMC article. Review.
-
Use of the Aplysia feeding network to study repetition priming of an episodic behavior.J Neurophysiol. 2017 Sep 1;118(3):1861-1870. doi: 10.1152/jn.00373.2017. Epub 2017 Jul 5. J Neurophysiol. 2017. PMID: 28679841 Free PMC article. Review.
References
-
- Alexeeva V, Borovikov D, Miller MW, Rosen SC, Cropper EC. Effect of a serotonergic extrinsic modulatory neuron (MCC) on radula mechanoafferent function in Aplysia. J Neurophysiol. 1998;80:1609–1622. - PubMed
-
- Arbas EA, Meinertzhagen IA, Shaw SR. Evolution in nervous systems. Annu Rev Neurosci. 1991;14:9–38. - PubMed
-
- Arnett BC (1996) Feeding and regurgitation: two modes of operation of the buccal central pattern generator in Helisoma PhD thesis, Chicago, IL: University of Illinois at Chicago.
-
- Arshavsky YI, Deliagina TG, Meizerov ES, Orlovsky GN, Panchin YV. Control of feeding movements in the freshwater snail, Planorbis corneus. I. Rhythmic neurons of buccal ganglia. Exp Brain Res. 1988;70:310–322. - PubMed
-
- Audesirk T, Audesirk G. Behavior of gastropod mollusks. In: Willows AOD, editor. The Mollusca. Neurobiology and Behavior, Part 1. Vol 8. Orlando, FL: Academic Press; 1985. pp. 2–94.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
