

Mechanisms by which human DNA primase chooses to polymerize a nucleoside triphosphate
- PMID: 20030400
- PMCID: PMC2847881
- DOI: 10.1021/bi9019516
Mechanisms by which human DNA primase chooses to polymerize a nucleoside triphosphate
Abstract
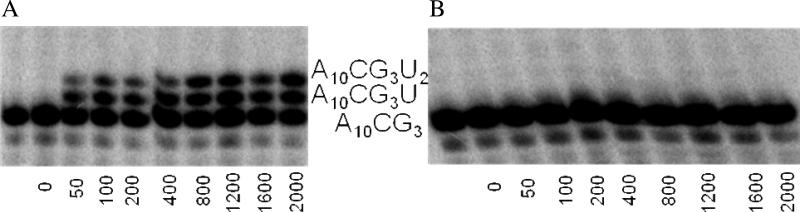
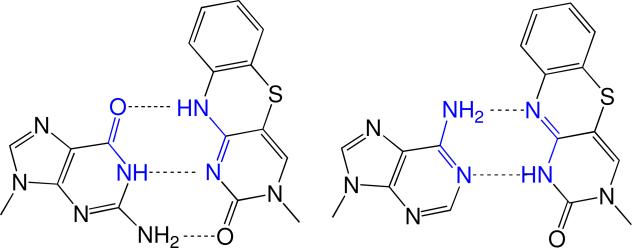
Human DNA primase synthesizes short RNA primers that DNA polymerase alpha then elongates during the initiation of all new DNA strands. Even though primase misincorporates NTPs at a relatively high frequency, this likely does not impact the final DNA product since the RNA primer is replaced with DNA. We used an extensive series of purine and pyrimidine analogues to provide further insights into the mechanism by which primase chooses whether or not to polymerize a NTP. Primase readily polymerized a size-expanded cytosine analogue, 1,3-diaza-2-oxophenothiazine NTP, across from a templating G but not across from A. The enzyme did not efficiently polymerize NTPs incapable of forming two Watson-Crick hydrogen bonds with the templating base with the exception of UTP opposite purine deoxyribonucleoside. Likewise, primase did not generate base pairs between two nucleotides with altered Watson-Crick hydrogen-bonding patterns. Examining the mechanism of NTP polymerization revealed that human primase can misincorporate NTPs via both template misreading and a primer-template slippage mechanism. Together, these data demonstrate that human primase strongly depends on Watson-Crick hydrogen bonds for efficient nucleotide polymerization, much more so than the mechanistically related herpes primase, and provide insights into the potential roles of primer-template stability and base tautomerization during misincorporation.
Figures




Similar articles
-
Herpes simplex virus-1 DNA primase: a remarkably inaccurate yet selective polymerase.Biochemistry. 2009 Nov 24;48(46):10866-81. doi: 10.1021/bi901476k. Biochemistry. 2009. PMID: 19835416 Free PMC article.
-
Human DNA primase uses Watson-Crick hydrogen bonds to distinguish between correct and incorrect nucleoside triphosphates.Biochemistry. 2004 Sep 28;43(38):12367-74. doi: 10.1021/bi0490791. Biochemistry. 2004. PMID: 15379576
-
Herpes simplex virus 1 primase employs watson-crick hydrogen bonding to identify cognate nucleoside triphosphates.Biochemistry. 2005 Nov 29;44(47):15585-93. doi: 10.1021/bi0513711. Biochemistry. 2005. PMID: 16300408
-
The Pol α-primase complex.Subcell Biochem. 2012;62:157-69. doi: 10.1007/978-94-007-4572-8_9. Subcell Biochem. 2012. PMID: 22918585 Review.
-
Mechanism and evolution of DNA primases.Biochim Biophys Acta. 2010 May;1804(5):1180-9. doi: 10.1016/j.bbapap.2009.06.011. Epub 2009 Jun 21. Biochim Biophys Acta. 2010. PMID: 19540940 Free PMC article. Review.
Cited by
-
Mechanism of Concerted RNA-DNA Primer Synthesis by the Human Primosome.J Biol Chem. 2016 May 6;291(19):10006-20. doi: 10.1074/jbc.M116.717405. Epub 2016 Mar 14. J Biol Chem. 2016. PMID: 26975377 Free PMC article.
-
The monomeric archaeal primase from Nanoarchaeum equitans harbours the features of heterodimeric archaeoeukaryotic primases and primes sequence-specifically.Nucleic Acids Res. 2023 Jun 9;51(10):5087-5105. doi: 10.1093/nar/gkad261. Nucleic Acids Res. 2023. PMID: 37099378 Free PMC article.
-
Divalent Cations Alter the Rate-Limiting Step of PrimPol-Catalyzed DNA Elongation.J Mol Biol. 2019 Feb 15;431(4):673-686. doi: 10.1016/j.jmb.2019.01.002. Epub 2019 Jan 8. J Mol Biol. 2019. PMID: 30633872 Free PMC article.
-
Insight into the Human DNA Primase Interaction with Template-Primer.J Biol Chem. 2016 Feb 26;291(9):4793-802. doi: 10.1074/jbc.M115.704064. Epub 2015 Dec 28. J Biol Chem. 2016. PMID: 26710848 Free PMC article.
-
Elaborated Action of the Human Primosome.Genes (Basel). 2017 Feb 8;8(2):62. doi: 10.3390/genes8020062. Genes (Basel). 2017. PMID: 28208743 Free PMC article. Review.
References
-
- Arezi B, Kuchta RD. Eukaryotic DNA Primase. Trends Biochem. Sci. 2000;25:572–576. - PubMed
-
- Kornberg A, Baker T. DNA Replication. 2 ed. W. H. Freeman & Co.; San Francisco: 1992.
-
- Pavlov YI, Frahm C, NickMcElhinny SA, Niimi A, Suzuki M, Kunkel TA. Evidence That Errors Made by DNA Polymerase alpha Are Corrected by DNA Polymerase delta. Curr. Biol. 2006;16:202–207. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
