

An examination of the proteolytic activity for bovine pregnancy-associated glycoproteins 2 and 12
- PMID: 20030586
- PMCID: PMC2969838
- DOI: 10.1515/bc.2010.016
An examination of the proteolytic activity for bovine pregnancy-associated glycoproteins 2 and 12
Abstract
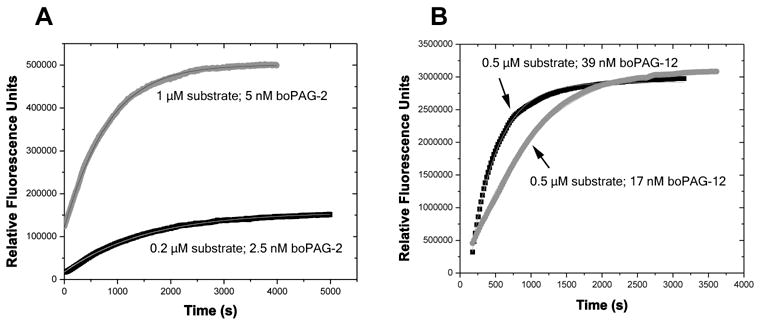
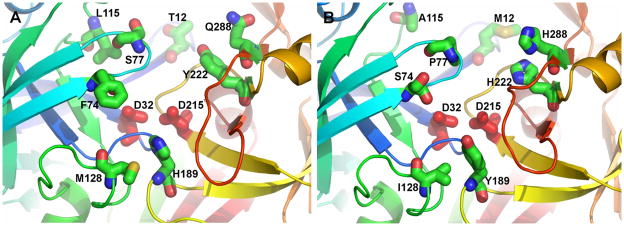
The pregnancy-associated glycoproteins (PAGs) represent a complex group of putative aspartic peptidases expressed exclusively in the placentas of species in the Artiodactyla order. The ruminant PAGs segregate into two classes: the 'ancient' and 'modern' PAGs. Some of the modern PAGs possess alterations in the catalytic center that are predicted to preclude their ability to act as peptidases. The ancient ruminant PAGs in contrast are thought to be peptidases, although no proteolytic activity has been described for these members. The aim of the present study was to investigate (1) if the ancient bovine PAGs (PAG-2 and PAG-12) have proteolytic activity, and (2) if there are any differences in activity between these two closely related members. Recombinant bovine PAG-2 and PAG-12 were expressed in a baculovirus expression system and the purified proteins were analyzed for proteolytic activity against a synthetic fluorescent cathepsin D/E substrate. Both proteins exhibited proteolytic activity with acidic pH optima. The k(cat)/K(m) for bovine PAG-2 was 2.7x10(5) m(-1) s(-1) and for boPAG-12 it was 6.8x10(4) m(-1) s(-1). The enzymes were inhibited by pepstatin A with a K(i) of 0.56 and 7.5 nm for boPAG-2 and boPAG-12, respectively. This is the first report describing proteolytic activity in PAGs from ruminant ungulates.
Figures
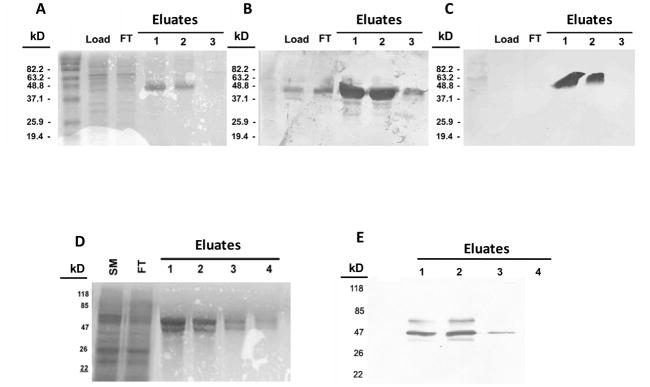
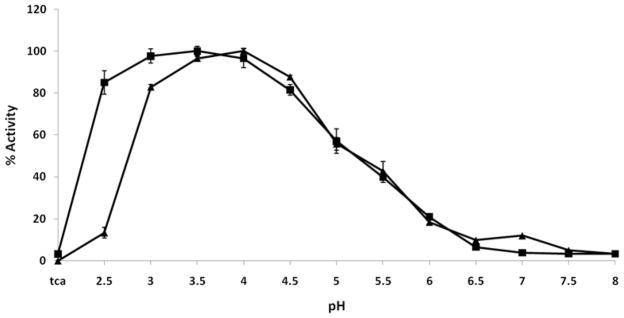
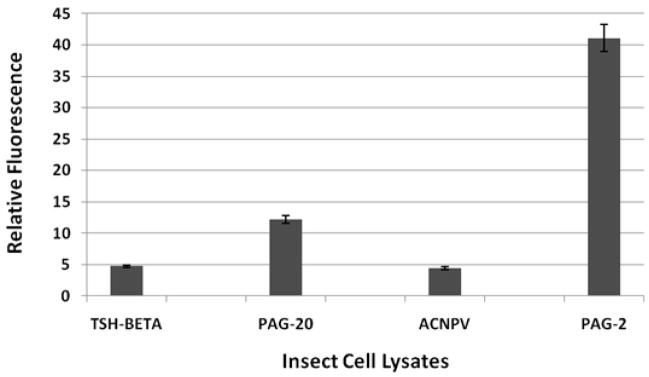
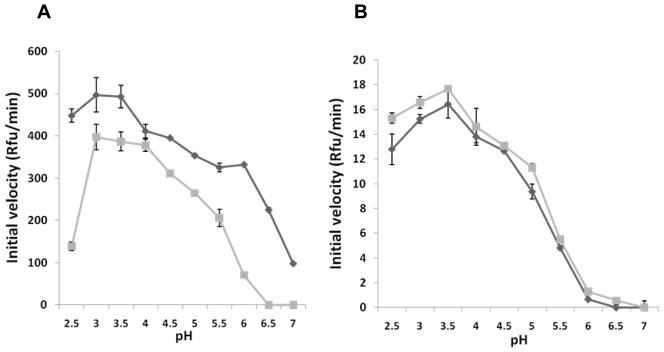
Similar articles
-
Characterization of the bovine pregnancy-associated glycoprotein gene family--analysis of gene sequences, regulatory regions within the promoter and expression of selected genes.BMC Genomics. 2009 Apr 24;10:185. doi: 10.1186/1471-2164-10-185. BMC Genomics. 2009. PMID: 19393060 Free PMC article.
-
Comparative modelling and analysis of amino acid substitutions suggests that the family of pregnancy-associated glycoproteins includes both active and inactive aspartic proteinases.Protein Eng. 1996 Oct;9(10):849-56. doi: 10.1093/protein/9.10.849. Protein Eng. 1996. PMID: 8931124
-
Characterization of the peptidase activity of recombinant porcine pregnancy-associated glycoprotein-2.J Biochem. 2008 Dec;144(6):725-32. doi: 10.1093/jb/mvn127. Epub 2008 Oct 3. J Biochem. 2008. PMID: 18835827
-
Biodiversity of multiple Pregnancy-Associated Glycoprotein (PAG) family: gene cloning and chorionic protein purification in domestic and wild eutherians (Placentalia)--a review.Reprod Nutr Dev. 2006 Sep-Oct;46(5):481-502. doi: 10.1051/rnd:2006034. Epub 2006 Sep 23. Reprod Nutr Dev. 2006. PMID: 17107639 Review.
-
Placental PAGs: gene origins, expression patterns, and use as markers of pregnancy.Reproduction. 2015 Mar;149(3):R115-26. doi: 10.1530/REP-14-0485. Reproduction. 2015. PMID: 25661256 Review.
Cited by
-
A multiplexed parallel reaction monitoring assay to monitor bovine pregnancy-associated glycoproteins throughout pregnancy and after gestation.PLoS One. 2022 Sep 23;17(9):e0271057. doi: 10.1371/journal.pone.0271057. eCollection 2022. PLoS One. 2022. PMID: 36149860 Free PMC article.
-
Effects of conceptus proteins on endometrium and blood leukocytes of dairy cattle using transcriptome and meta-analysis.bioRxiv [Preprint]. 2024 Apr 28:2024.04.25.591148. doi: 10.1101/2024.04.25.591148. bioRxiv. 2024. PMID: 38712302 Free PMC article. Preprint.
-
mRNA expression profiling in cotyledons reveals significant up-regulation of the two bovine pregnancy-associated glycoprotein genes boPAG-8 and boPAG-11 in early gestation.Vet Med Sci. 2018 Nov;4(4):341-350. doi: 10.1002/vms3.123. Epub 2018 Sep 28. Vet Med Sci. 2018. PMID: 30265452 Free PMC article.
-
Crystal structure of a putative aspartic proteinase domain of the Mycobacterium tuberculosis cell surface antigen PE_PGRS16.FEBS Open Bio. 2013 Jun 8;3:256-62. doi: 10.1016/j.fob.2013.05.004. Print 2013. FEBS Open Bio. 2013. PMID: 23923105 Free PMC article.
-
Primary structures of different isoforms of buffalo pregnancy-associated glycoproteins (BuPAGs) during early pregnancy and elucidation of the 3-dimensional structure of the most abundant isoform BuPAG 7.PLoS One. 2018 Nov 7;13(11):e0206143. doi: 10.1371/journal.pone.0206143. eCollection 2018. PLoS One. 2018. PMID: 30403702 Free PMC article.
References
-
- Abad-Zapatero C, Rydel TJ, Erickson JW. Revised 2.3 A0 structure of porcine pepsin: evidence for a flexible subdomain. Proteins. 1990;8:62–71. - PubMed
-
- Arnold K, Bordoli L, Kopp J, Schwede T. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics. 2006;22:195–201. - PubMed
-
- Barrett A, Rawlings N, Woessner J. Hanbook of proteolytic enzymes. Elsevier Academic Press; San Diego: 2004.
-
- Beyer BB, Johnson JV, Chung AY, Li T, Madabushi A, Agbandje-McKenna M, McKenna R, Dame JB, et al. Active-Site Specificity of Digestive Aspartic Peptidases from the Four Species of Plasmodium that Infect Humans Using Chromogenic Combinatorial Peptide Libraries. Biochemistry. 2005;44:1768–1779. - PubMed
-
- Brandt GA, Parks TE, Killian G, Ealy AD, Green JA. A cloning and expression analysis of pregnancy-associated glycoproteins expressed in trophoblasts of the white-tail deer placenta. Mol Reprod Dev. 2007;74:1355–1362. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous