

Membrane-bound guaiacol peroxidases from maize (Zea mays L.) roots are regulated by methyl jasmonate, salicylic acid, and pathogen elicitors
- PMID: 20032108
- PMCID: PMC2814115
- DOI: 10.1093/jxb/erp353
Membrane-bound guaiacol peroxidases from maize (Zea mays L.) roots are regulated by methyl jasmonate, salicylic acid, and pathogen elicitors
Abstract
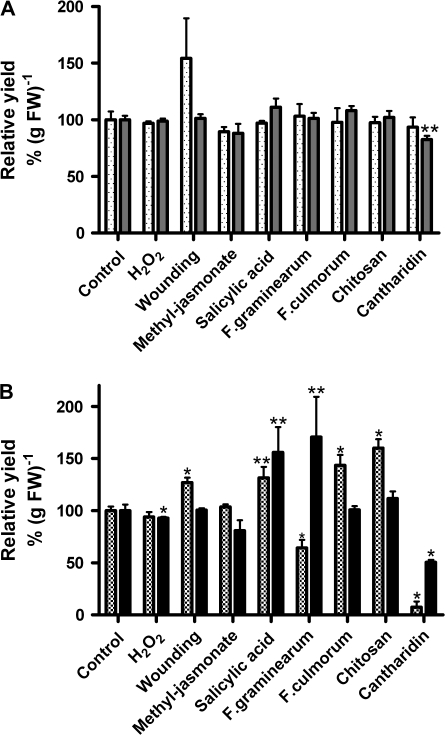
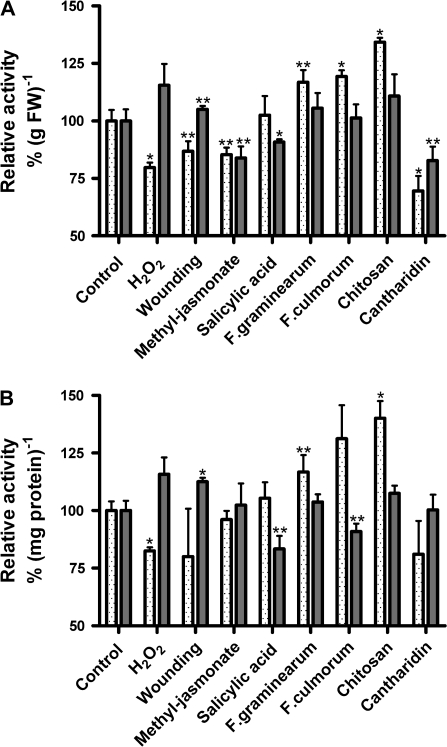
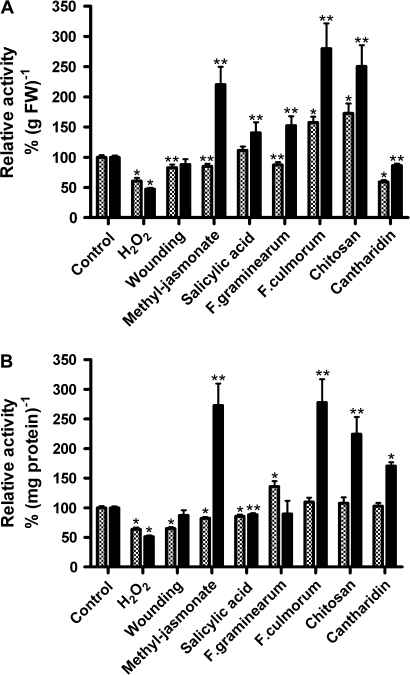
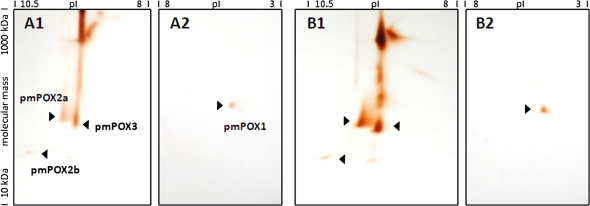
Plant peroxidases are involved in numerous cellular processes in plant development and stress responses. Four plasma membrane-bound peroxidases have been identified and characterized in maize (Zea mays L.) roots. In the present study, maize seedlings were treated with different stresses and signal compounds, and a functional analysis of these membrane-bound class III peroxidases (pmPOX1, pmPOX2a, pmPOX2b, and pmPOX3) was carried out. Total guaiacol peroxidase activities from soluble and microsomal fractions of maize roots were compared and showed weak changes. By contrast, total plasma membrane and washed plasma membrane peroxidase activities, representing peripheral and integral membrane proteins, revealed strong changes after all of the stresses applied. A proteomic approach using 2D-PAGE analysis showed that pmPOX3 was the most abundant class III peroxidase at plasma membranes of control plants, followed by pmPOX2a >pmPOX2b >pmPOX1. The molecular mass (63 kDa) and the isoelectric point (9.5) of the pmPOX2a monomer were identified for the first time. The protein levels of all four enzymes changed in response to multiple stresses. While pmPOX2b was the only membrane peroxidase down-regulated by wounding, all four enzymes were differentially but strongly stimulated by methyl jasmonate, salicylic acid, and elicitors (Fusarium graminearum and Fusarium culmorum extracts, and chitosan) indicating their function in pathogen defence. Oxidative stress applied as H(2)O(2) treatment up-regulated pmPOX2b >pmPOX2a, while pmPOX3 was down-regulated. Treatment with the phosphatase inhibitor chantharidin resulted in distinct responses.
Figures
Similar articles
-
Hypoxia-Responsive Class III Peroxidases in Maize Roots: Soluble and Membrane-Bound Isoenzymes.Int J Mol Sci. 2020 Nov 23;21(22):8872. doi: 10.3390/ijms21228872. Int J Mol Sci. 2020. PMID: 33238617 Free PMC article.
-
Membrane-bound class III peroxidases: identification, biochemical properties and sequence analysis of isoenzymes purified from maize (Zea mays L.) roots.J Proteomics. 2008 Oct 7;71(4):412-24. doi: 10.1016/j.jprot.2008.06.006. Epub 2008 Jun 17. J Proteomics. 2008. PMID: 18603027
-
Combined seed and foliar pre-treatments with exogenous methyl jasmonate and salicylic acid mitigate drought-induced stress in maize.PLoS One. 2020 May 1;15(5):e0232269. doi: 10.1371/journal.pone.0232269. eCollection 2020. PLoS One. 2020. PMID: 32357181 Free PMC article.
-
An integrated proteomic approach to decipher the effect of methyl jasmonate elicitation on the proteome of Silybum marianum L. hairy roots.Plant Physiol Biochem. 2013 Sep;70:115-22. doi: 10.1016/j.plaphy.2013.05.031. Epub 2013 May 31. Plant Physiol Biochem. 2013. PMID: 23771036
-
Membrane-Bound Class III Peroxidases: Unexpected Enzymes with Exciting Functions.Int J Mol Sci. 2018 Sep 21;19(10):2876. doi: 10.3390/ijms19102876. Int J Mol Sci. 2018. PMID: 30248965 Free PMC article. Review.
Cited by
-
Physiological basis of differential zinc and copper tolerance of Verbascum populations from metal-contaminated and uncontaminated areas.Environ Sci Pollut Res Int. 2016 May;23(10):10005-20. doi: 10.1007/s11356-016-6177-4. Epub 2016 Feb 11. Environ Sci Pollut Res Int. 2016. PMID: 26865485
-
Hypoxia-Responsive Class III Peroxidases in Maize Roots: Soluble and Membrane-Bound Isoenzymes.Int J Mol Sci. 2020 Nov 23;21(22):8872. doi: 10.3390/ijms21228872. Int J Mol Sci. 2020. PMID: 33238617 Free PMC article.
-
Genome-wide and evolutionary analysis of the class III peroxidase gene family in wheat and Aegilops tauschii reveals that some members are involved in stress responses.BMC Genomics. 2019 Aug 22;20(1):666. doi: 10.1186/s12864-019-6006-5. BMC Genomics. 2019. PMID: 31438842 Free PMC article.
-
Characterization of a gene encoding clathrin heavy chain in maize up-regulated by salicylic acid, abscisic acid and high boron supply.Int J Mol Sci. 2013 Jul 22;14(7):15179-98. doi: 10.3390/ijms140715179. Int J Mol Sci. 2013. PMID: 23880865 Free PMC article.
-
Pre-fractionation strategies to resolve pea (Pisum sativum) sub-proteomes.Front Plant Sci. 2015 Oct 19;6:849. doi: 10.3389/fpls.2015.00849. eCollection 2015. Front Plant Sci. 2015. PMID: 26539198 Free PMC article. Review.
References
-
- Almagro L, Gómez Ros LV, Belchi-Navarro S, Bru R, Ros Barceló A, Pedreño MA. Class III peroxidases in plant defence reactions. Journal of Experimental Botany. 2009;60:377–390. - PubMed
-
- Asada K, Miyake C, Ogawa K, Hossain MA. Microcompartmentation of ascorbate peroxidase and regeneration of ascorbate from ascorbate radical: its dual role in chloroplasts. In: Obinger C, Burner U, Ebermann R, Penel C, Greppin H, editors. Proceedings of the IV international symposium on plant peroxidases: biochemistry and physiolology. Switzerland: University of Vienna, Austria and University of Geneva; 1996. pp. 163–167.
-
- Bérczi A, Asard H. Soluble proteins, an often overlooked contaminant in plasma membrane preparations. Trends in Plant Science. 2003;8:250–251. - PubMed
-
- Bienert GP, Schjoerring JK, Jahn TP. Membrane transport of hydrogen peroxide. Biochimica et Biophysica Acta. 2006;1758:994–1003. - PubMed
-
- Blom N, Gammeltoft S, Brunak S. Sequence- and structure-based prediction of eukaryotic protein phosphorylation sites. Journal of Molecular Biology. 1999;294:1351–1362. - PubMed
