

Lesions of the auditory cortex impair azimuthal sound localization and its recalibration in ferrets
- PMID: 20032231
- PMCID: PMC2887622
- DOI: 10.1152/jn.00991.2009
Lesions of the auditory cortex impair azimuthal sound localization and its recalibration in ferrets
Abstract
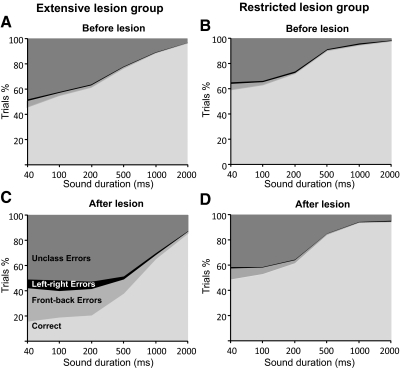
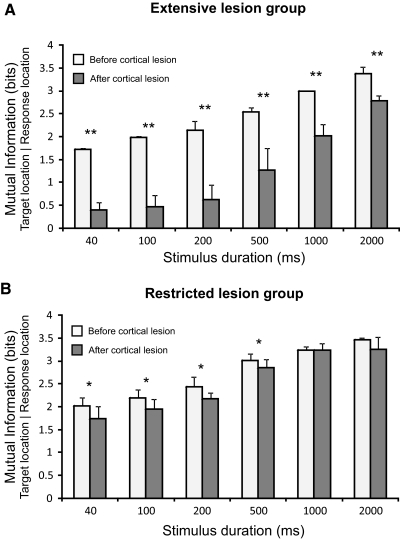
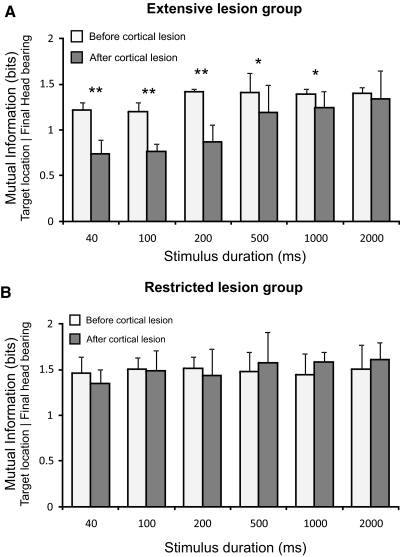
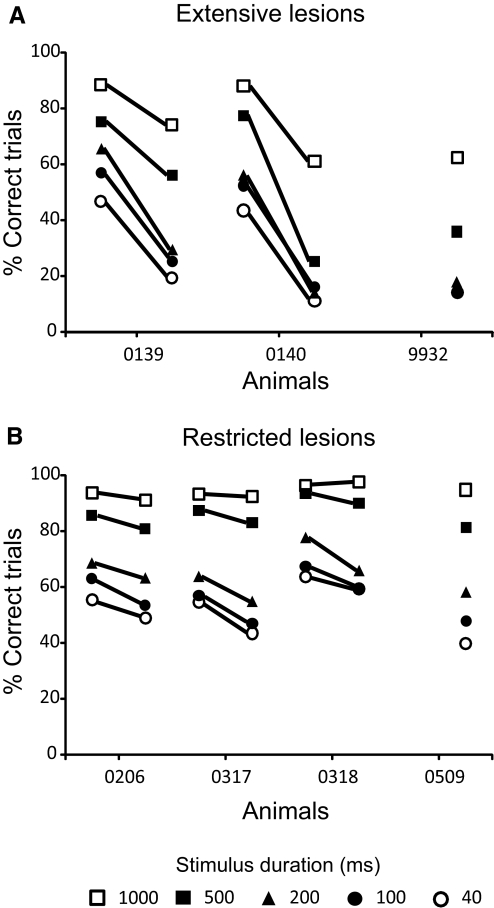
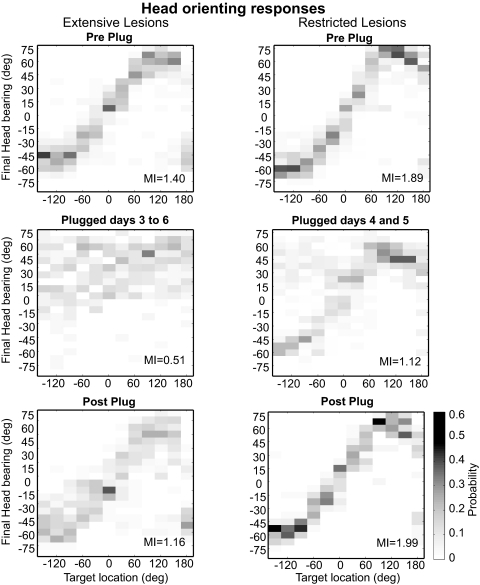
The role of auditory cortex in sound localization and its recalibration by experience was explored by measuring the accuracy with which ferrets turned toward and approached the source of broadband sounds in the horizontal plane. In one group, large bilateral lesions were made of the middle ectosylvian gyrus, where the primary auditory cortical fields are located, and part of the anterior and/or posterior ectosylvian gyrus, which contain higher-level fields. In the second group, the lesions were intended to be confined to primary auditory cortex (A1). The ability of the animals to localize noise bursts of different duration and level was measured before and after the lesions were made. A1 lesions produced a modest disruption of approach-to-target responses to short-duration stimuli (<500 ms) on both sides of space, whereas head orienting accuracy was unaffected. More extensive lesions produced much greater auditory localization deficits, again primarily for shorter sounds. In these ferrets, the accuracy of both the approach-to-target behavior and the orienting responses was impaired, and they could do little more than correctly lateralize the stimuli. Although both groups of ferrets were still able to localize long-duration sounds accurately, they were, in contrast to ferrets with an intact auditory cortex, unable to relearn to localize these stimuli after altering the spatial cues available by reversibly plugging one ear. These results indicate that both primary and nonprimary cortical areas are necessary for normal sound localization, although only higher auditory areas seem to contribute to accurate head orienting behavior. They also show that the auditory cortex, and A1 in particular, plays an essential role in training-induced plasticity in adult ferrets, and that this is the case for both head orienting responses and approach-to-target behavior.
Figures





Similar articles
-
Cortical cholinergic input is required for normal auditory perception and experience-dependent plasticity in adult ferrets.J Neurosci. 2013 Apr 10;33(15):6659-71. doi: 10.1523/JNEUROSCI.5039-12.2013. J Neurosci. 2013. PMID: 23575862 Free PMC article.
-
Physiological and behavioral studies of spatial coding in the auditory cortex.Hear Res. 2007 Jul;229(1-2):106-15. doi: 10.1016/j.heares.2007.01.001. Epub 2007 Jan 17. Hear Res. 2007. PMID: 17314017 Free PMC article. Review.
-
Sound localization during homotopic and heterotopic bilateral cooling deactivation of primary and nonprimary auditory cortical areas in the cat.J Neurophysiol. 2007 Jan;97(1):26-43. doi: 10.1152/jn.00720.2006. Epub 2006 Oct 11. J Neurophysiol. 2007. PMID: 17035367
-
Sound localization deficits during reversible deactivation of primary auditory cortex and/or the dorsal zone.J Neurophysiol. 2008 Apr;99(4):1628-42. doi: 10.1152/jn.01228.2007. Epub 2008 Jan 16. J Neurophysiol. 2008. PMID: 18199813
-
[The role of the auditory cortex in the spatial information processing].Rev Neurol. 2012 Jul 16;55(2):91-100. Rev Neurol. 2012. PMID: 22760769 Review. Spanish.
Cited by
-
Development of Questionnaire for Auditory Localization.J Int Adv Otol. 2018 Aug;14(2):197-201. doi: 10.5152/iao.2018.4243. J Int Adv Otol. 2018. PMID: 30256196 Free PMC article.
-
Integrating information from different senses in the auditory cortex.Biol Cybern. 2012 Dec;106(11-12):617-25. doi: 10.1007/s00422-012-0502-x. Epub 2012 Jul 14. Biol Cybern. 2012. PMID: 22798035 Free PMC article. Review.
-
Cortical cholinergic input is required for normal auditory perception and experience-dependent plasticity in adult ferrets.J Neurosci. 2013 Apr 10;33(15):6659-71. doi: 10.1523/JNEUROSCI.5039-12.2013. J Neurosci. 2013. PMID: 23575862 Free PMC article.
-
Auditory gap-in-noise detection behavior in ferrets and humans.Behav Neurosci. 2015 Aug;129(4):473-90. doi: 10.1037/bne0000065. Epub 2015 Jun 8. Behav Neurosci. 2015. PMID: 26052794 Free PMC article.
-
The non-lemniscal auditory cortex in ferrets: convergence of corticotectal inputs in the superior colliculus.Front Neuroanat. 2010 May 21;4:18. doi: 10.3389/fnana.2010.00018. eCollection 2010. Front Neuroanat. 2010. PMID: 20640247 Free PMC article.
References
-
- Beitel RE, Kaas JH. Effects of bilateral and unilateral ablation of auditory cortex in cats on the unconditioned head orienting response to acoustic stimuli. J Neurophysiol 70: 351–369, 1993 - PubMed
-
- Bizley JK, Nodal FR, Nelken I, King AJ. Functional organization of ferret auditory cortex. Cereb Cortex 15: 1637–1653, 2005 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources