

Plasticity-related gene 5 (PRG5) induces filopodia and neurite growth and impedes lysophosphatidic acid- and nogo-A-mediated axonal retraction
- PMID: 20032306
- PMCID: PMC2820418
- DOI: 10.1091/mbc.e09-06-0506
Plasticity-related gene 5 (PRG5) induces filopodia and neurite growth and impedes lysophosphatidic acid- and nogo-A-mediated axonal retraction
Abstract
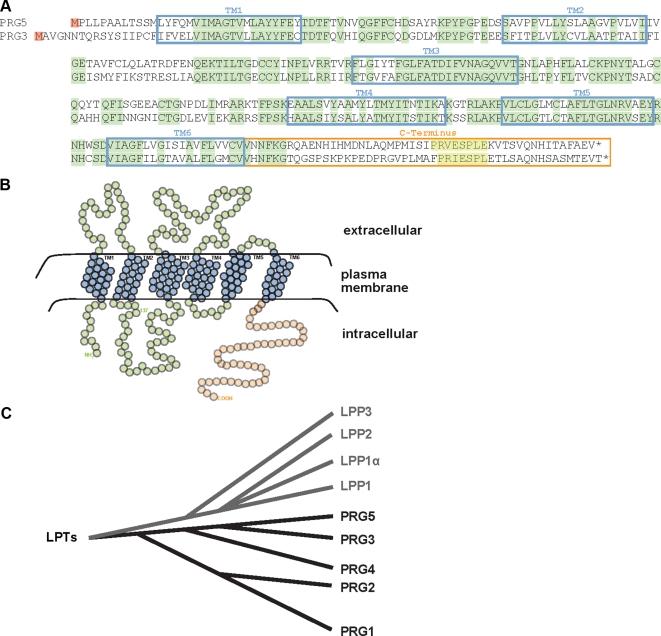
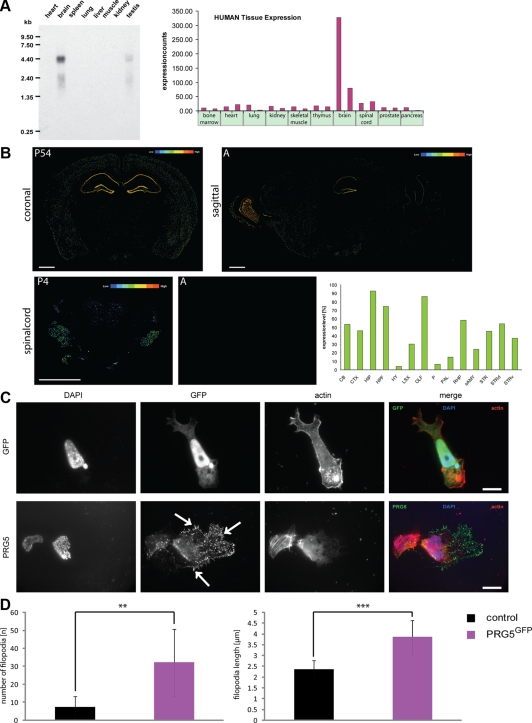
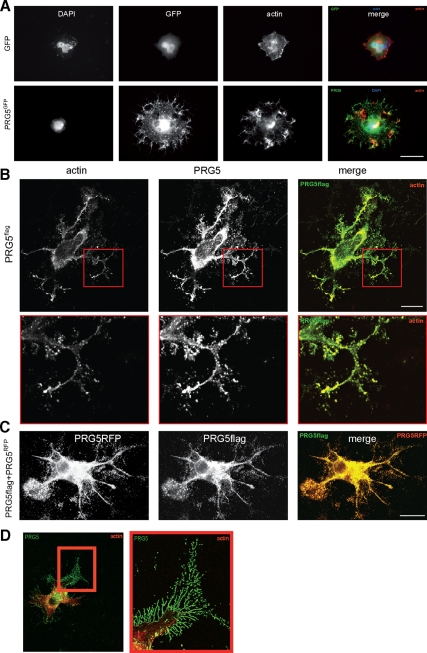
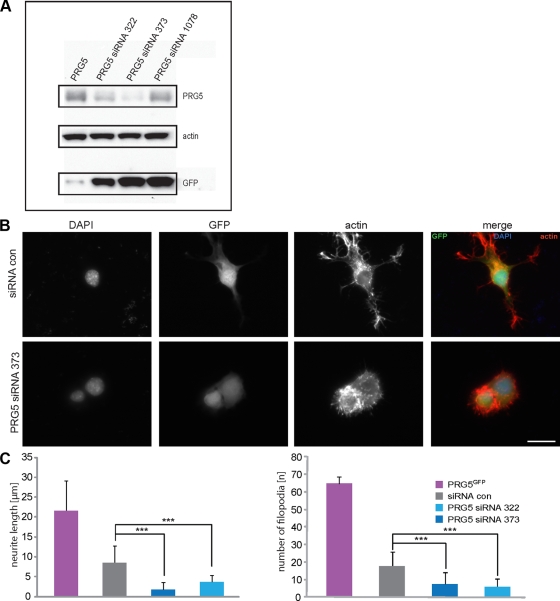
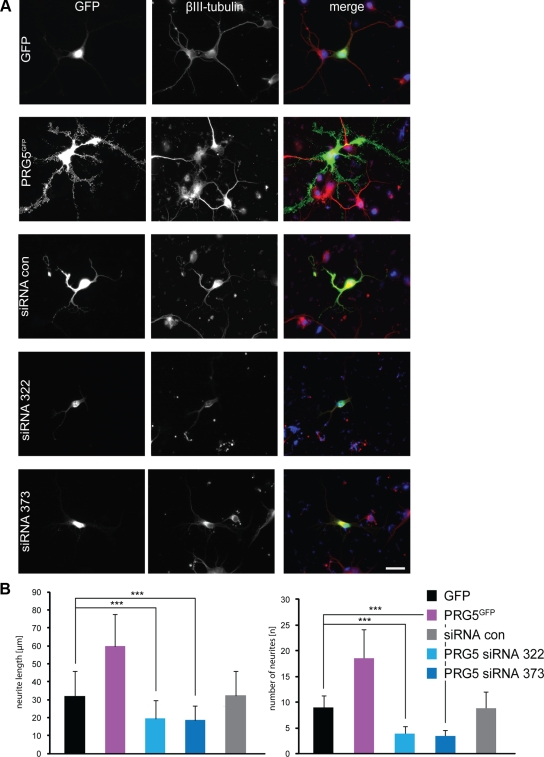
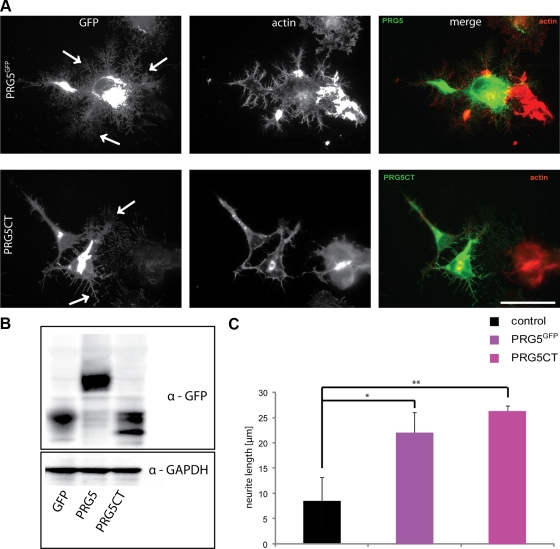
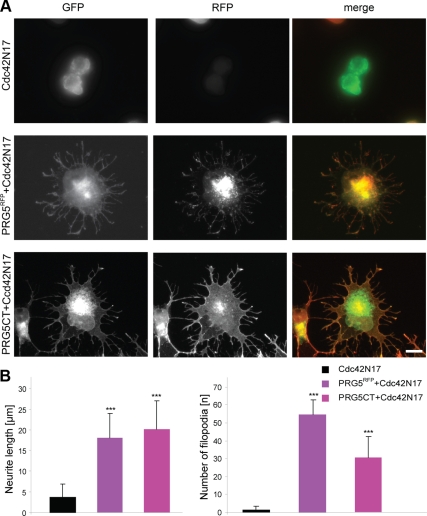
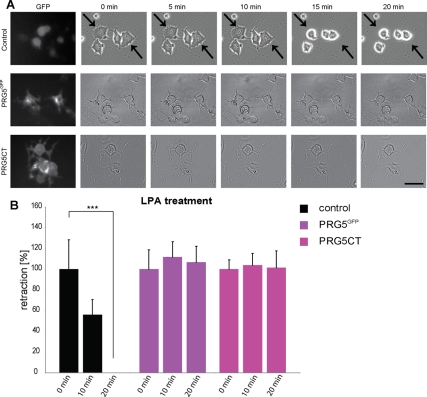
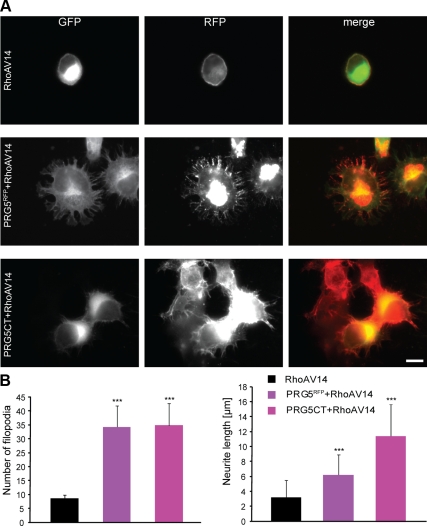
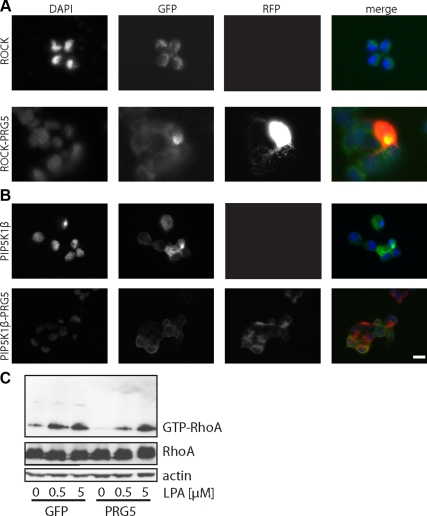
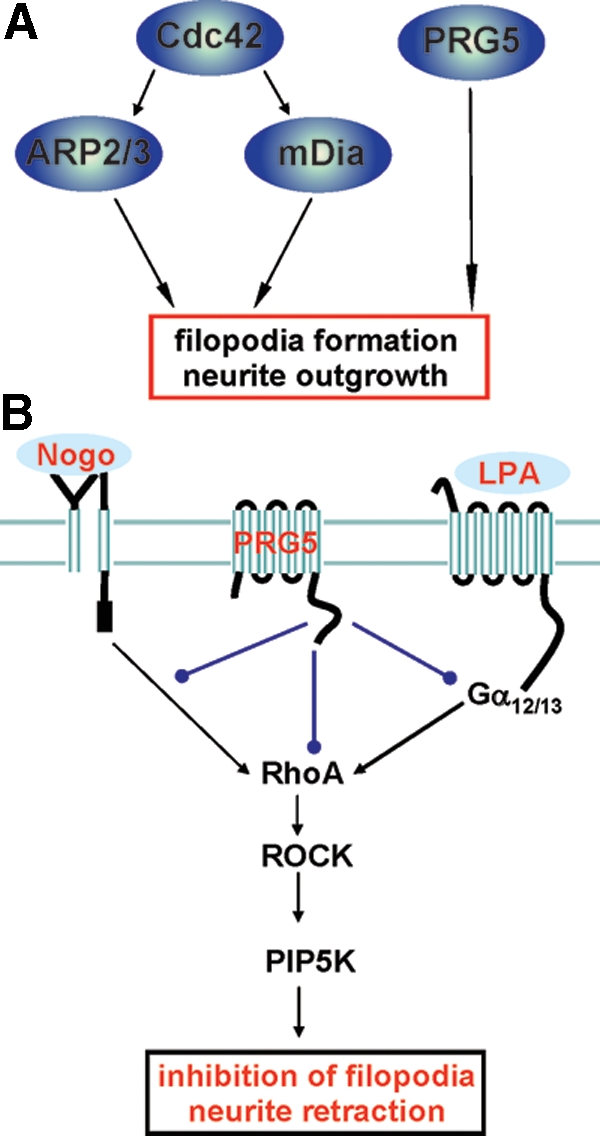
Members of the plasticity-related gene (PRG1-4) family are brain-specific integral membrane proteins and implicated in neuronal plasticity, such as filopodia formation and axon growth after brain lesion. Here we report on the cloning of a novel member of the PRG family, PRG5, with high homologies to PRG3. PRG5 is regulated during brain and spinal cord development and is exclusively allocated within the nervous system. When introduced in neurons, PRG5 is distributed in the plasma membrane and induces filopodia as well as axon elongation and growth. Conversely, siRNA mediated knockdown of PRG5 impedes axon growth and disturbs filopodia formation. Here we show that PRG5 induces filopodia growth independently of Cdc42. Moreover, axon collapse and RhoA activation induced by LPA and myelin-associated neurite inhibitor Nogo-A is attenuated in the presence of PRG5, although direct activation of the RhoA-Rho-PIP5K kinase pathway abolishes PRG5 -formed neurites. Thus, we describe here the identification of a novel member of the PRG family that induces filopodia and axon elongation in a Cdc42-independent manner. In addition, PRG5 impedes brain injury-associated growth inhibitory signals upstream of the RhoA-Rho kinase pathway.
Figures

References
-
- Abe T., Kato M., Miki H., Takenawa T., Endo T. Small GTPase Tc10 and its homologue RhoT induce N-WASP-mediated long process formation and neurite outgrowth. J. Cell Sci. 2003;116:155–168. - PubMed
-
- Atwal J. K., Pinkston-Gosse J., Syken J., Stawicki S., Wu Y., Shatz C., Tessier-Lavigne M. PirB is a functional receptor for myelin inhibitors of axonal regeneration. Science. 2008;322:967–970. - PubMed
-
- Benfey M., Aguayo A. J. Extensive elongation of axons from rat brain into peripheral nerve graft. Nature. 1982;296:150–152. - PubMed
-
- Bräuer A. U., Savaskan N. E., Kühn H., Prehn S., Ninnemann O., Nitsch R. A new phospholipid phosphatase, PRG-1, is involved in axon growth and regenerative sprouting. Nat. Neurosci. 2003;6:572–578. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
