

Autophosphorylation of polo-like kinase 4 and its role in centriole duplication
- PMID: 20032307
- PMCID: PMC2820420
- DOI: 10.1091/mbc.e09-06-0505
Autophosphorylation of polo-like kinase 4 and its role in centriole duplication
Abstract
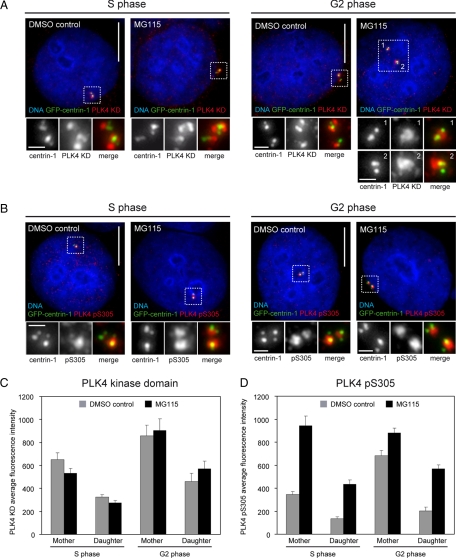
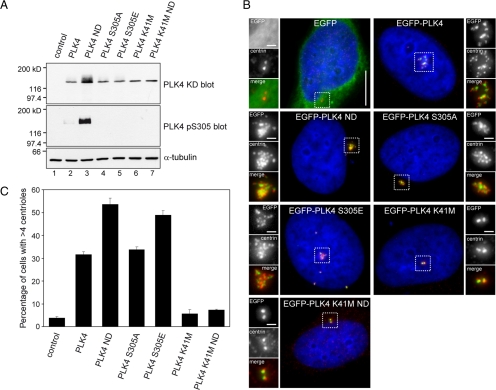
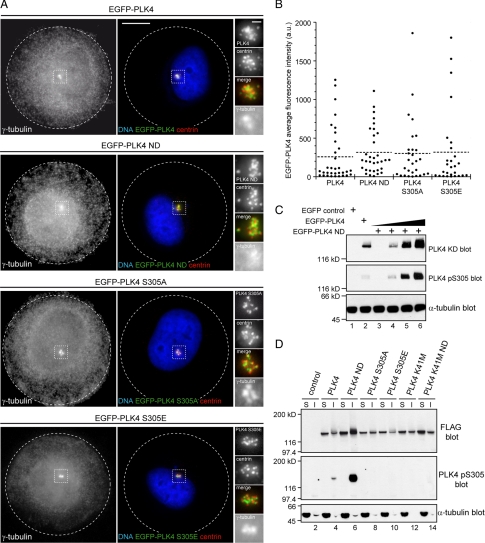
Centrosome duplication occurs once every cell cycle in a strictly controlled manner. Polo-like kinase 4 (PLK4) is a key regulator of this process whose kinase activity is essential for centriole duplication. Here, we show that PLK4 autophosphorylation of serine S305 is a consequence of kinase activation and enables the active fraction to be identified in the cell. Active PLK4 is detectable on the replicating mother centriole in G1/S, with the proportion of active kinase increasing through interphase to reach a maximum in mitosis. Activation of PLK4 at the replicating daughter centriole is delayed until G2, but a level equivalent to the replicating mother centriole is achieved in M phase. Active PLK4 is regulated by the proteasome, because either proteasome inhibition or mutation of the degron motif of PLK4 results in the accumulation of S305-phosphorylated PLK4. Autophosphorylation probably plays a role in the process of centriole duplication, because mimicking S305 phosphorylation enhances the ability of overexpressed PLK4 to induce centriole amplification. Importantly, we show that S305-phosphorylated PLK4 is specifically sequestered at the centrosome contrary to the nonphosphorylated form. These data suggest that PLK4 activity is restricted to the centrosome to prevent aberrant centriole assembly and sustained kinase activity is required for centriole duplication.
Figures

Similar articles
-
Cep152 interacts with Plk4 and is required for centriole duplication.J Cell Biol. 2010 Nov 15;191(4):721-9. doi: 10.1083/jcb.201006049. Epub 2010 Nov 8. J Cell Biol. 2010. PMID: 21059850 Free PMC article.
-
Plk4 trans-autophosphorylation regulates centriole number by controlling betaTrCP-mediated degradation.J Cell Sci. 2010 Jul 1;123(Pt 13):2163-9. doi: 10.1242/jcs.068502. Epub 2010 Jun 1. J Cell Sci. 2010. PMID: 20516151
-
Cep152 acts as a scaffold for recruitment of Plk4 and CPAP to the centrosome.J Cell Biol. 2010 Nov 15;191(4):731-9. doi: 10.1083/jcb.201007107. Epub 2010 Nov 8. J Cell Biol. 2010. PMID: 21059844 Free PMC article.
-
Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication-Impact on Cancer.Cells. 2022 Feb 24;11(5):786. doi: 10.3390/cells11050786. Cells. 2022. PMID: 35269408 Free PMC article. Review.
-
The PLK4-STIL-SAS-6 module at the core of centriole duplication.Biochem Soc Trans. 2016 Oct 15;44(5):1253-1263. doi: 10.1042/BST20160116. Biochem Soc Trans. 2016. PMID: 27911707 Free PMC article. Review.
Cited by
-
A novel Cep120-dependent mechanism inhibits centriole maturation in quiescent cells.Elife. 2018 May 9;7:e35439. doi: 10.7554/eLife.35439. Elife. 2018. PMID: 29741480 Free PMC article.
-
TRIM37 prevents formation of centriolar protein assemblies by regulating Centrobin.Elife. 2021 Jan 25;10:e62640. doi: 10.7554/eLife.62640. Elife. 2021. PMID: 33491649 Free PMC article.
-
KAT2A/KAT2B-targeted acetylome reveals a role for PLK4 acetylation in preventing centrosome amplification.Nat Commun. 2016 Oct 31;7:13227. doi: 10.1038/ncomms13227. Nat Commun. 2016. PMID: 27796307 Free PMC article.
-
Primary ciliogenesis requires the distal appendage component Cep123.Biol Open. 2013 Apr 9;2(6):535-45. doi: 10.1242/bio.20134457. Print 2013 Jun 15. Biol Open. 2013. PMID: 23789104 Free PMC article.
-
Expression of Polo-Like Kinase 4(PLK4) in Breast Cancer and Its Response to Taxane-Based Neoadjuvant Chemotherapy.J Cancer. 2016 Jun 6;7(9):1125-32. doi: 10.7150/jca.14307. eCollection 2016. J Cancer. 2016. PMID: 27326256 Free PMC article.
References
-
- Azioune A., Storch M., Bornens M., Thery M., Piel M. Simple and rapid process for single cell micro-patterning. Lab Chip. 2009;9:1640–1642. - PubMed
-
- Bettencourt-Dias M., Rodrigues-Martins A., Carpenter L., Riparbelli M., Lehmann L., Gatt M. K., Carmo N., Balloux F., Callaini G., Glover D. M. SAK/PLK4 is required for centriole duplication and flagella development. Curr. Biol. 2005;15:2199–2207. - PubMed
-
- Bornens M. Centrosome composition and microtubule anchoring mechanisms. Curr. Opin. Cell Biol. 2002;14:25–34. - PubMed
-
- Chen Z., Indjeian V. B., McManus M., Wang L., Dynlacht B. D. CP110, a cell cycle-dependent CDK substrate, regulates centrosome duplication in human cells. Dev. Cell. 2002;3:339–350. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
