

Structure of an integrin with an alphaI domain, complement receptor type 4
- PMID: 20033057
- PMCID: PMC2830704
- DOI: 10.1038/emboj.2009.367
Structure of an integrin with an alphaI domain, complement receptor type 4
Abstract
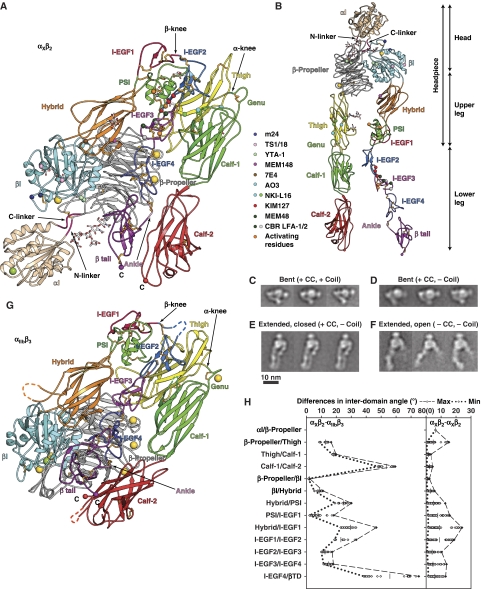
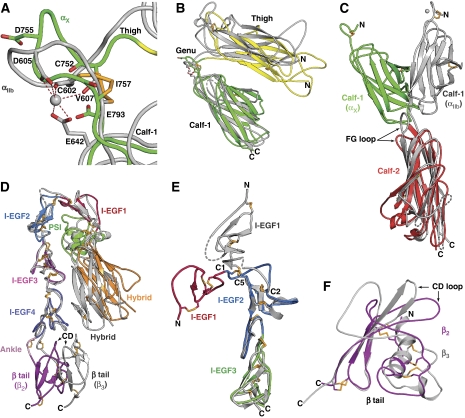
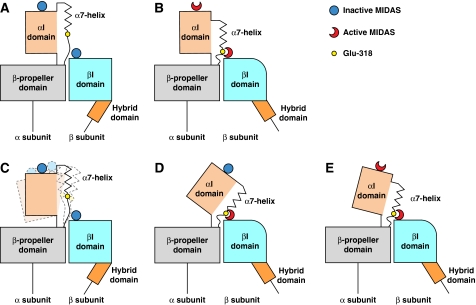
We report the structure of an integrin with an alphaI domain, alpha(X)beta(2), the complement receptor type 4. It was earlier expected that a fixed orientation between the alphaI domain and the beta-propeller domain in which it is inserted would be required for allosteric signal transmission. However, the alphaI domain is highly flexible, enabling two betaI domain conformational states to couple to three alphaI domain states, and greater accessibility for ligand recognition. Although alpha(X)beta(2) is bent similarly to integrins that lack alphaI domains, the terminal domains of the alpha- and beta-legs, calf-2 and beta-tail, are oriented differently than in alphaI-less integrins. Linkers extending to the transmembrane domains are unstructured. Previous mutations in the beta(2)-tail domain support the importance of extension, rather than a deadbolt, in integrin activation. The locations of further activating mutations and antibody epitopes show the critical role of extension, and conversion from the closed to the open headpiece conformation, in integrin activation. Differences among 10 molecules in crystal lattices provide unprecedented information on interdomain flexibility important for modelling integrin extension and activation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC (2002) PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr 58: 1948–1954 - PubMed
-
- Alonso JL, Essafi M, Xiong JP, Stehle T, Arnaout MA (2002) Does the integrin αA domain act as a ligand for its βA domain? Curr Biol 12: R340–R342 - PubMed
-
- Bailey S (1994) The CCP4 suite-programs for protein crystallography. Acta Crystallogr 50: 760–763 - PubMed
-
- Beglova N, Blacklow SC, Takagi J, Springer TA (2002) Cysteine-rich module structure reveals a fulcrum for integrin rearrangement upon activation. Nat Struct Biol 9: 282–287 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
