

The evolution of postpollination reproductive isolation in Costus
- PMID: 20033446
- PMCID: PMC2796120
- DOI: 10.1007/s00497-009-0113-4
The evolution of postpollination reproductive isolation in Costus
Abstract
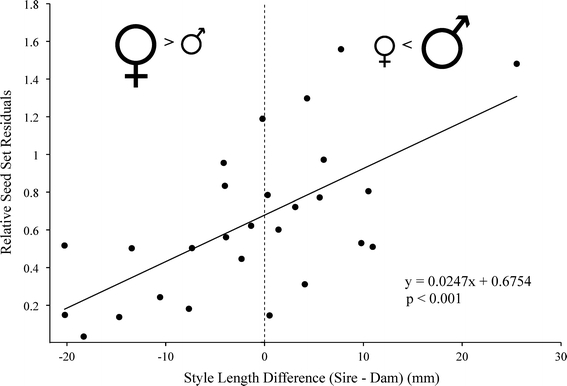
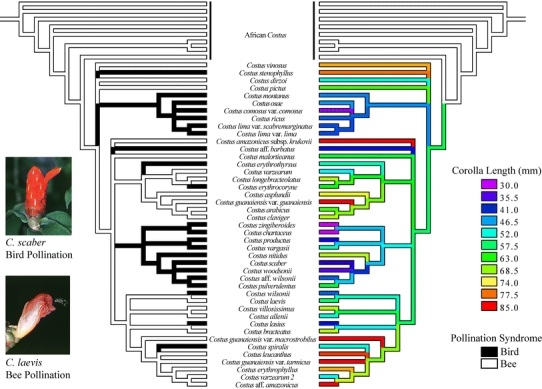
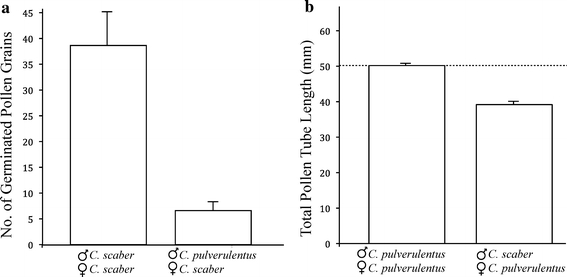
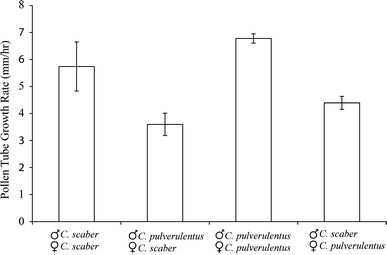
Reproductive isolation is critical to the diversification of species. Postpollination barriers may be important in limiting gene flow between closely related species, but they are relatively cryptic and their evolution is poorly understood. Here, we review the role of postpollination reproductive isolation in plants, including the various stages at which it operates and the hypotheses for how it may evolve. We then review empirical studies in the plant genus Costus, evaluating documented postpollination barriers in light of these hypotheses. We summarize isolation due to parental style length differences and present evidence supporting the hypothesis that the differences are in part a by-product of selection on floral morphology. Additionally, we show that reduced pollen adhesion, germination, and tube growth contribute to reproductive isolation between two closely related sympatric species of Costus. Geographic variation in the strength of these crossing barriers supports the hypothesis that they evolved under reinforcement, or direct natural selection to strengthen isolation.
Keywords: Costus; Postpollination; Reinforcement; Reproductive isolation.
Figures

Similar articles
-
Reproductive isolation between two closely related hummingbird-pollinated neotropical gingers.Evolution. 2006 Mar;60(3):538-52. Evolution. 2006. PMID: 16637499
-
Floral isolation is the main reproductive barrier among closely related sexually deceptive orchids.Evolution. 2011 Sep;65(9):2606-20. doi: 10.1111/j.1558-5646.2011.01323.x. Epub 2011 Jun 9. Evolution. 2011. PMID: 21884059
-
Natural selection reinforces speciation in a radiation of neotropical rainforest plants.Evolution. 2008 Oct;62(10):2628-42. doi: 10.1111/j.1558-5646.2008.00463.x. Epub 2008 Sep 18. Evolution. 2008. PMID: 18637960
-
The origins of reproductive isolation in plants.New Phytol. 2015 Sep;207(4):968-84. doi: 10.1111/nph.13424. Epub 2015 May 5. New Phytol. 2015. PMID: 25944305 Review.
-
Speciation in Daphnia.Mol Ecol. 2021 Mar;30(6):1398-1418. doi: 10.1111/mec.15824. Epub 2021 Mar 1. Mol Ecol. 2021. PMID: 33522056 Review.
Cited by
-
The Evolution of Sex is Tempered by Costly Hybridization in Boechera (Rock Cress).J Hered. 2021 Mar 12;112(1):67-77. doi: 10.1093/jhered/esaa041. J Hered. 2021. PMID: 33211850 Free PMC article.
-
The timetable for allopolyploidy in flowering plants.Ann Bot. 2013 Nov;112(7):1201-8. doi: 10.1093/aob/mct194. Epub 2013 Aug 21. Ann Bot. 2013. PMID: 23965616 Free PMC article. Review.
-
Mismatch between pollen and pistil size causes asymmetric mechanical reproductive isolation across Phlox species.bioRxiv [Preprint]. 2024 May 10:2024.05.08.593106. doi: 10.1101/2024.05.08.593106. bioRxiv. 2024. Update in: Evolution. 2024 Dec 2;78(12):1936-1948. doi: 10.1093/evolut/qpae128. PMID: 38766021 Free PMC article. Updated. Preprint.
-
On the Coyne and Orr-igin of species: effects of intrinsic postzygotic isolation, ecological differentiation, x chromosome size, and sympatry on Drosophila speciation.Evolution. 2014 Apr;68(4):1176-87. doi: 10.1111/evo.12330. Epub 2014 Jan 26. Evolution. 2014. PMID: 24325145 Free PMC article.
-
Eighteen new species of Neotropical Costaceae (Zingiberales).PhytoKeys. 2023 Mar 22;222:75-127. doi: 10.3897/phytokeys.222.87779. eCollection 2023. PhytoKeys. 2023. PMID: 37252638 Free PMC article.
References
-
- Arnold ML, Hamrick JL, Bennett BD. Interspecific pollen competition and reproductive isolation in Iris. J Hered. 1993;84:13.
-
- Bella JL, Butlin RK, Ferris C, Hewitt GM. Asymmetrical homogamy and unequal sex ratio from reciprocal mating-order crosses between Chorthippus parallelus subspecies. Heredity. 1992;68:345–352.
-
- Carney SE, Hodges SA, Arnold ML. Effects of differential pollen-tube growth on hybridization in the Louisiana irises. Evol Int J org Evol. 1996;50:1871–1878. - PubMed
