

Synaptic development of the mouse dorsal lateral geniculate nucleus
- PMID: 20034053
- PMCID: PMC4278806
- DOI: 10.1002/cne.22223
Synaptic development of the mouse dorsal lateral geniculate nucleus
Abstract
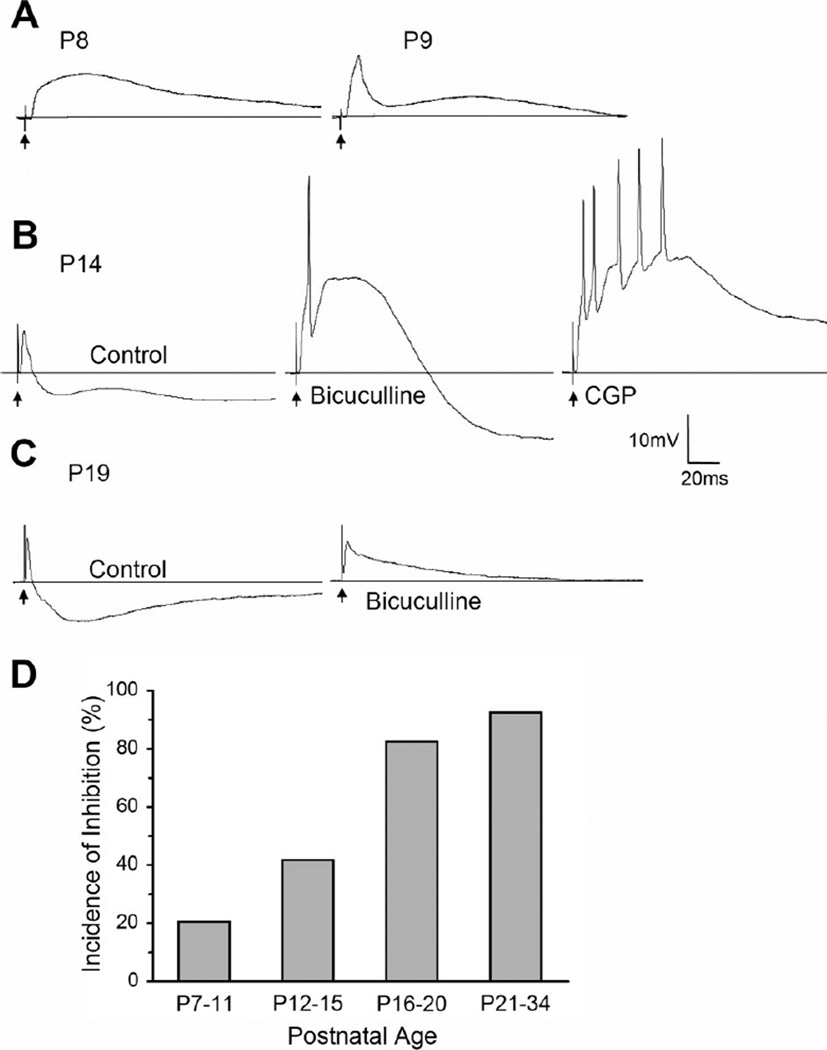
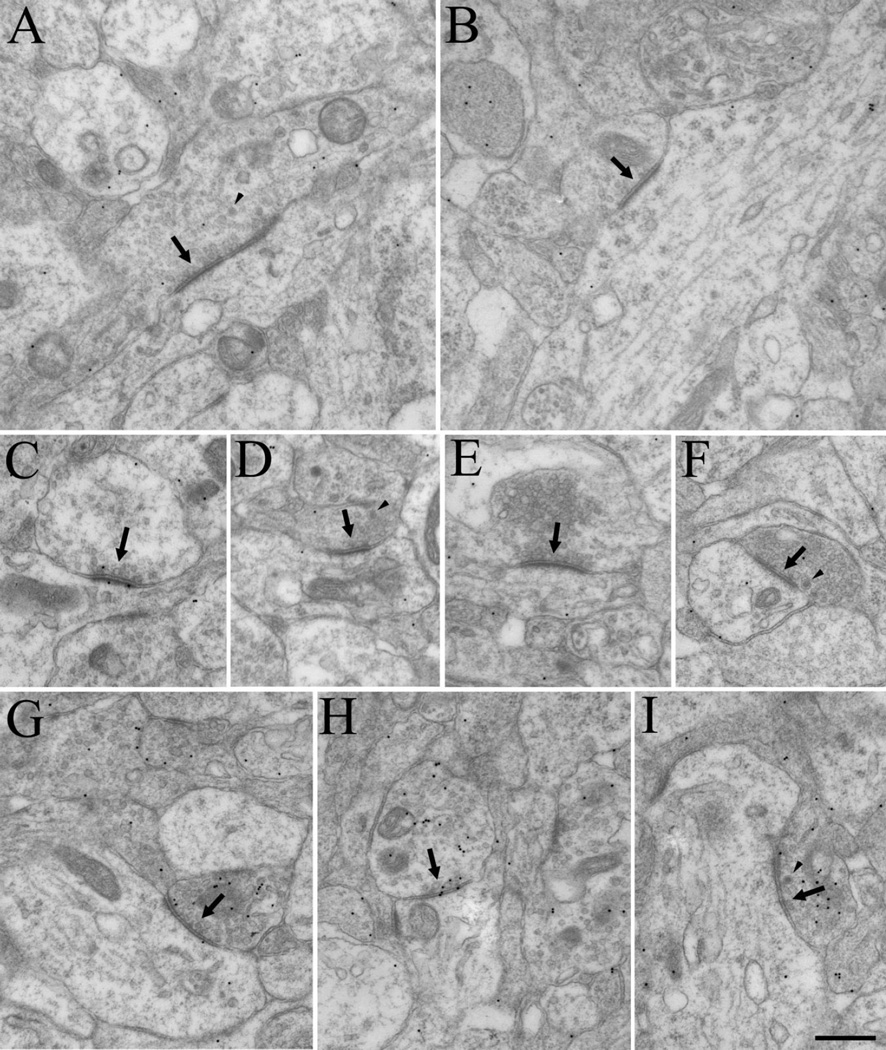
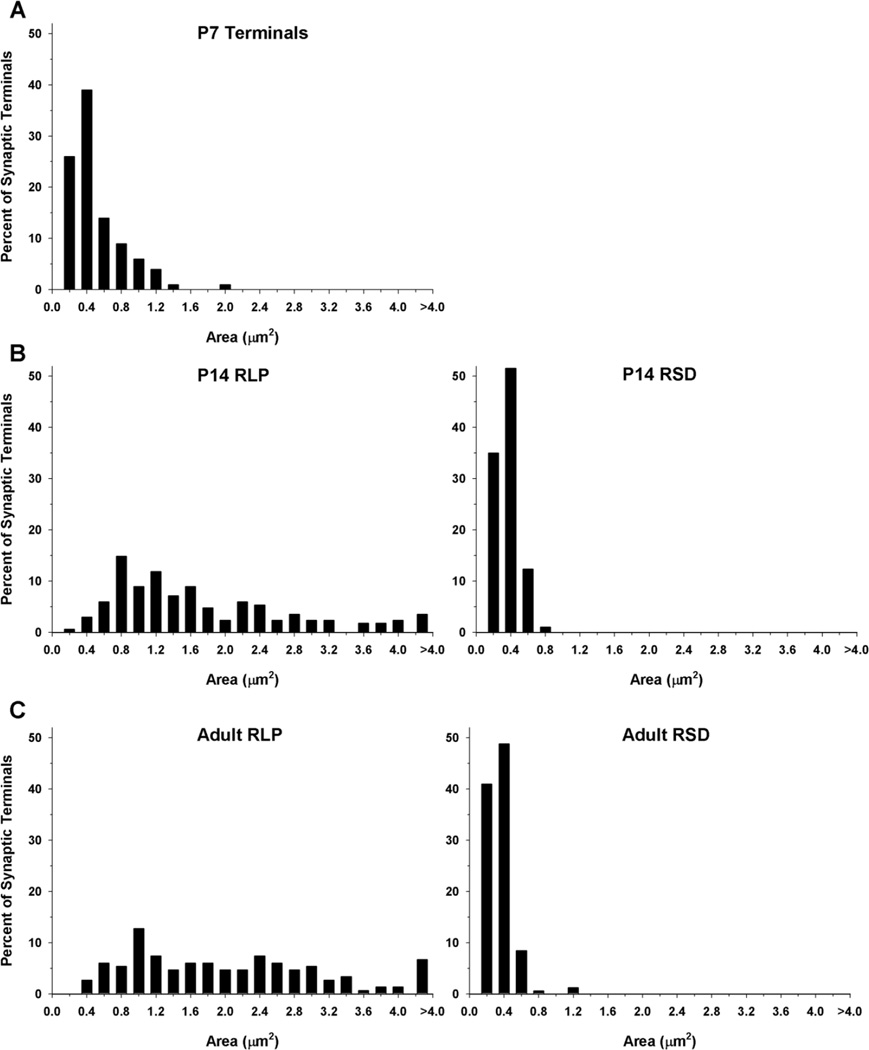
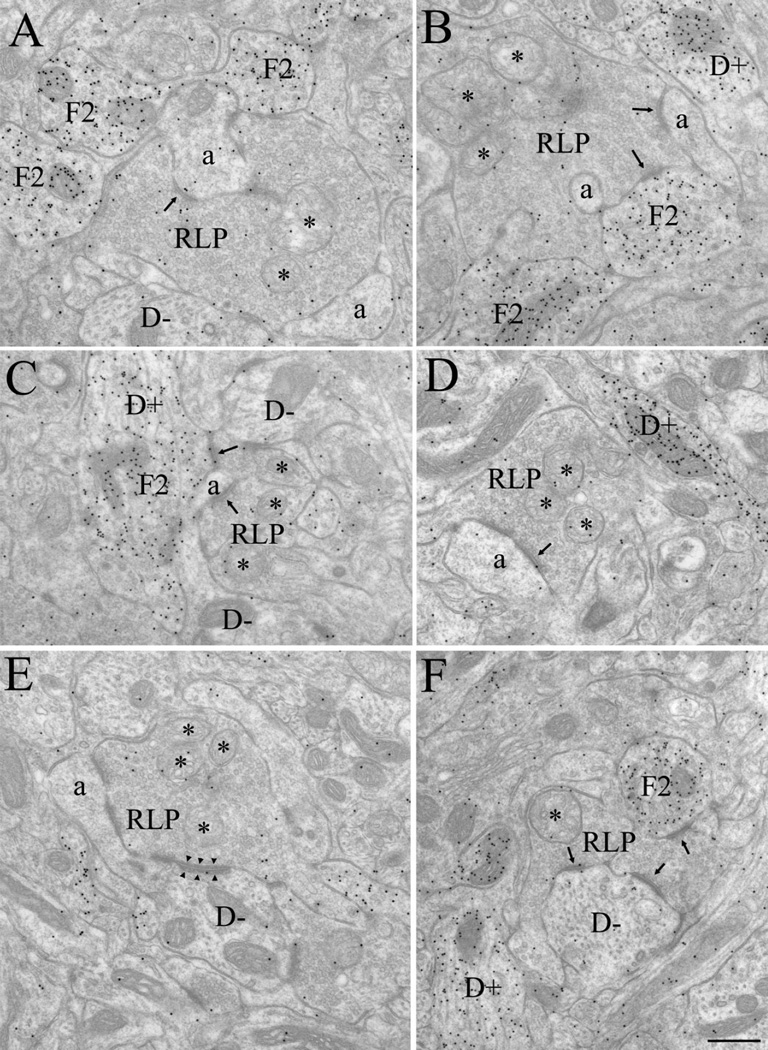
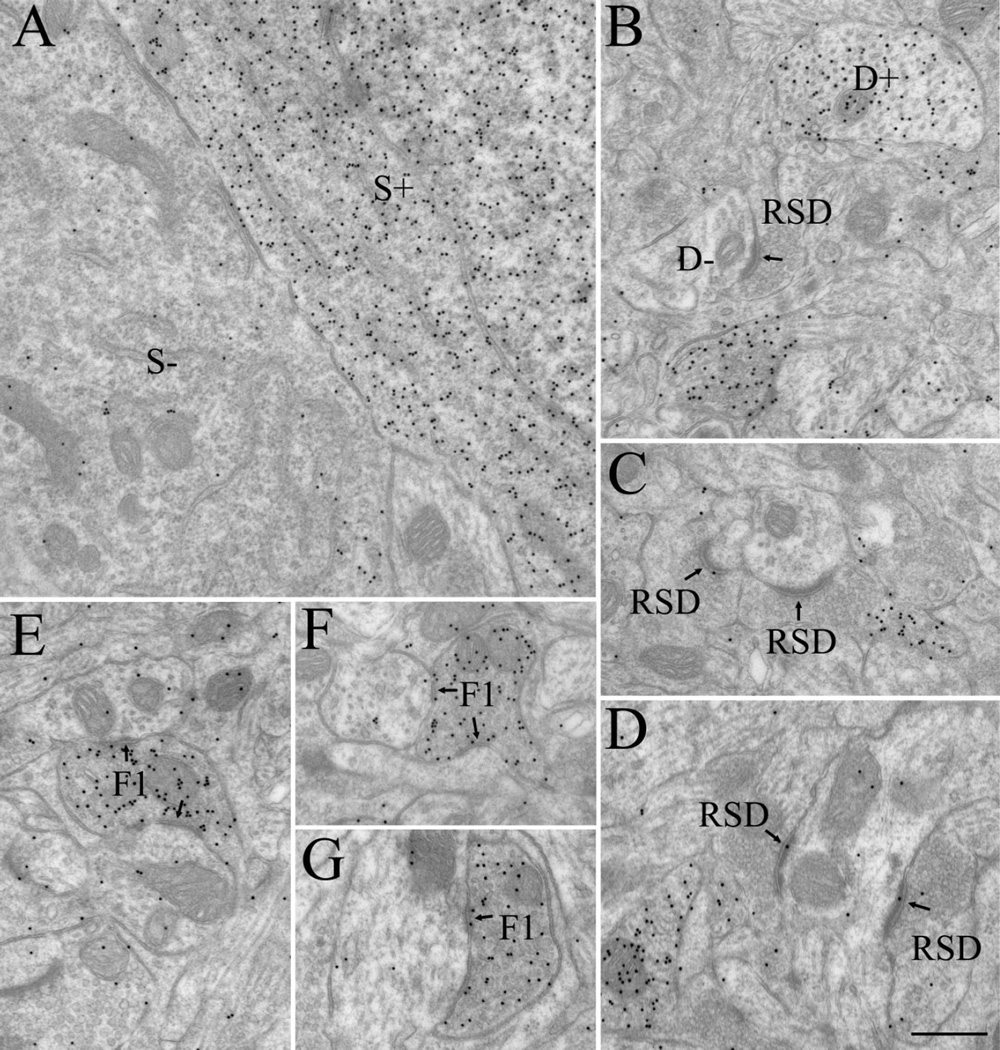
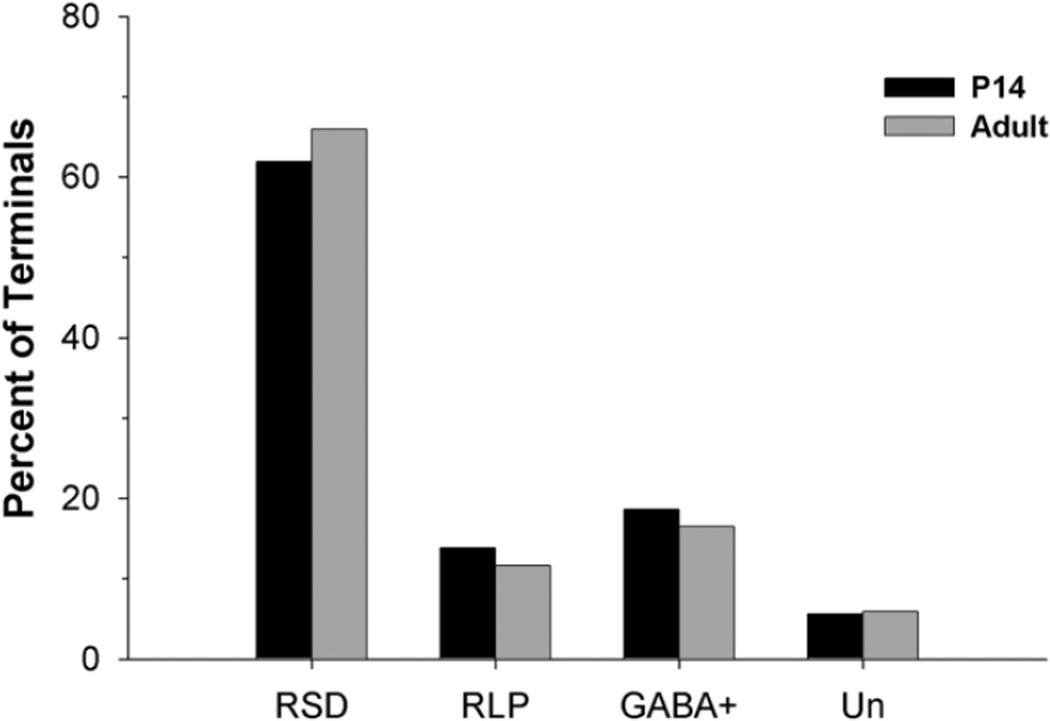
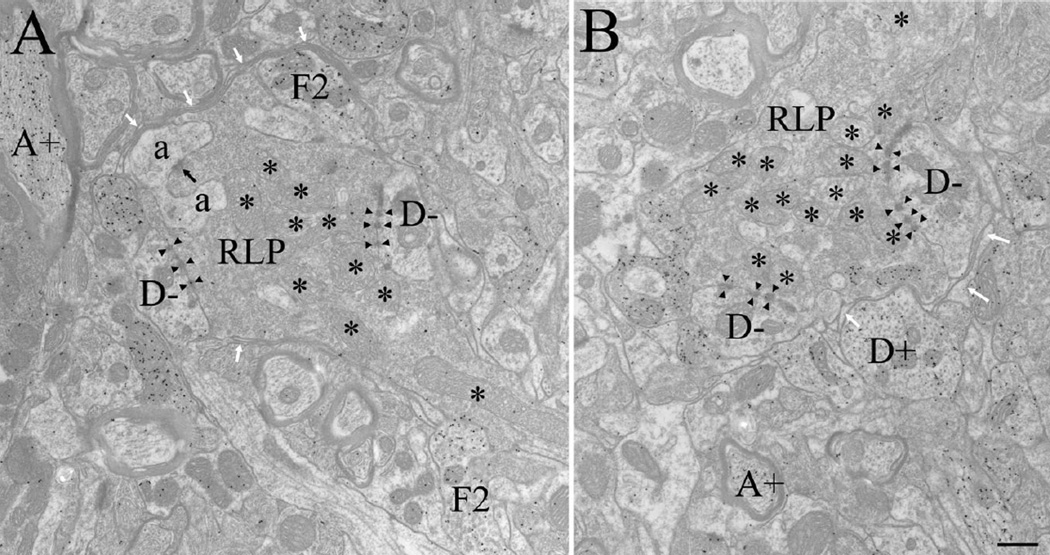
The dorsal lateral geniculate nucleus (dLGN) of the mouse has emerged as a model system in the study of thalamic circuit development. However, there is still a lack of information regarding how and when various types of retinal and nonretinal synapses develop. We examined the synaptic organization of the developing mouse dLGN in the common pigmented C57/BL6 strain, by recording the synaptic responses evoked by electrical stimulation of optic tract axons, and by investigating the ultrastructure of identified synapses. At early postnatal ages (<P12), optic tract evoked responses were primarily excitatory. The full complement of inhibitory responses did not emerge until after eye opening (>P14), when optic tract stimulation routinely evoked an excitatory postsynaptic potential/inhibitory postsynaptic potential (EPSP/IPSP) sequence, with the latter having both a GABA(A) and GABA(B) component. Electrophysiological and ultrastructural observations were consistent. At P7, many synapses were present, but synaptic profiles lacked the ultrastructural features characteristic of the adult dLGN, and little gamma-aminobutyric acid (GABA) could be detected by using immunocytochemical techniques. In contrast, by P14, GABA staining was robust, mature synaptic profiles of retinal and nonretinal origin were easily distinguished, and the size and proportion of synaptic contacts were similar to those of the adult. The emergence of nonretinal synapses coincides with pruning of retinogeniculate connections, and the transition of retinal activity from spontaneous to visually driven. These results indicate that the synaptic architecture of the mouse dLGN is similar to that of other higher mammals, and thus provides further support for its use as a model system for visual system development.
2009 Wiley-Liss, Inc.
Figures

References
-
- Arcelli P, Frassoni C, Regondi MC, De Biasi S, Spreafico R. GABAergic neurons in mammalian thalamus: a marker of thalamic complexity? Brain Res Bull. 1997;42:27–37. - PubMed
-
- Bentivoglio M, Spreafico R, Alvarez-Bolado G, Sanchez MP, Fairen A. Differential expression of the GABAA receptor complex in the dorsal thalamus and reticular nucleus: an immunohistochemical study in the adult and developing rat. Eur J Neurosci. 1991;3:118–125. - PubMed
-
- Blitz DM, Regehr WG. Timing and specificity of feed-forward inhibition within the LGN. Neuron. 2005;45:917–928. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
