

Churchill and Sip1a repress fibroblast growth factor signaling during zebrafish somitogenesis
- PMID: 20034103
- PMCID: PMC2962425
- DOI: 10.1002/dvdy.22201
Churchill and Sip1a repress fibroblast growth factor signaling during zebrafish somitogenesis
Abstract
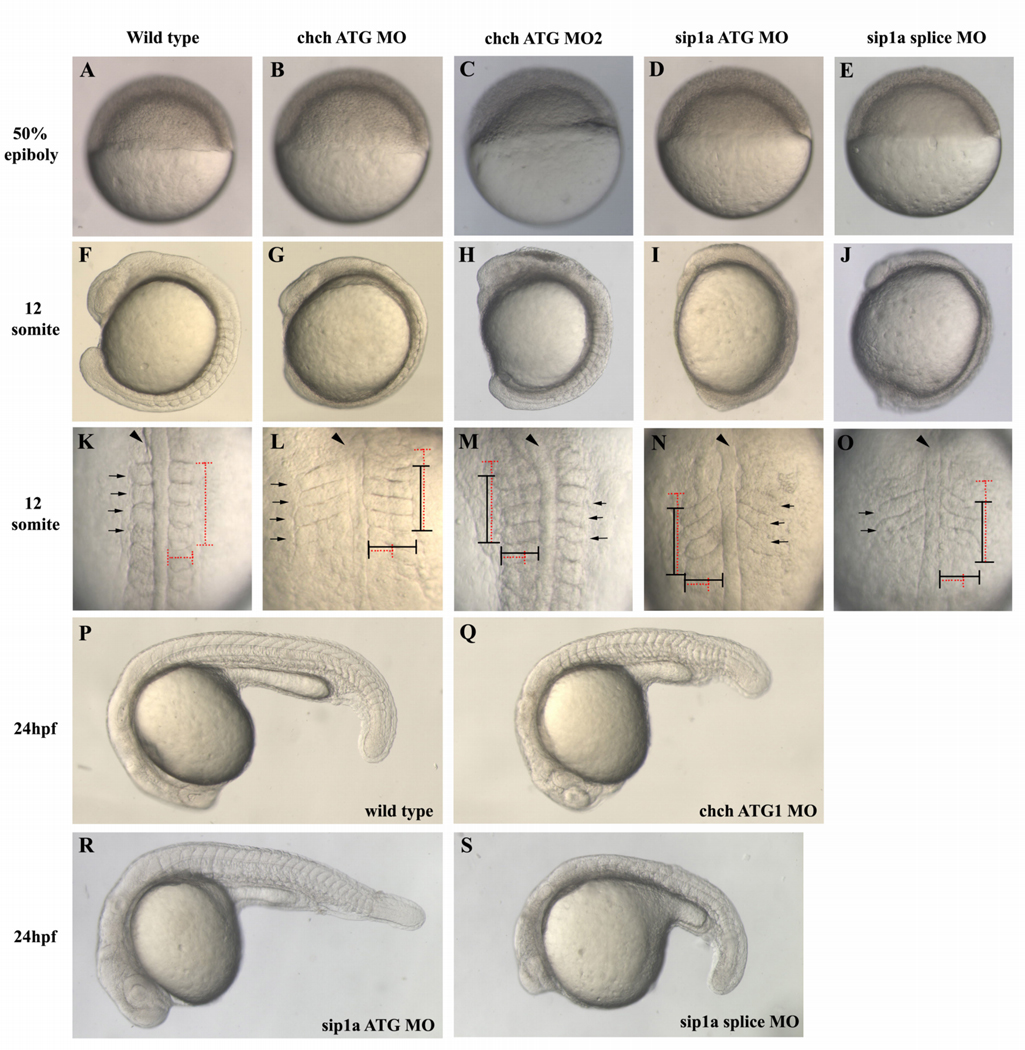
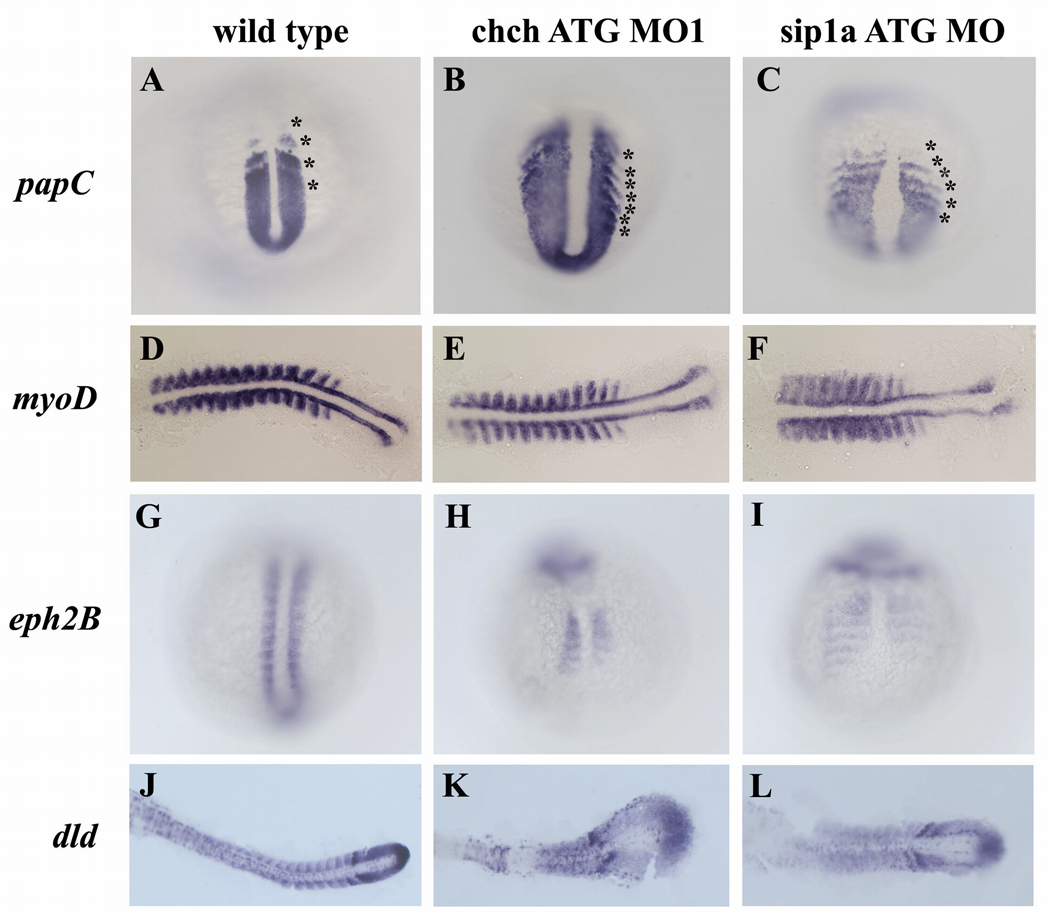
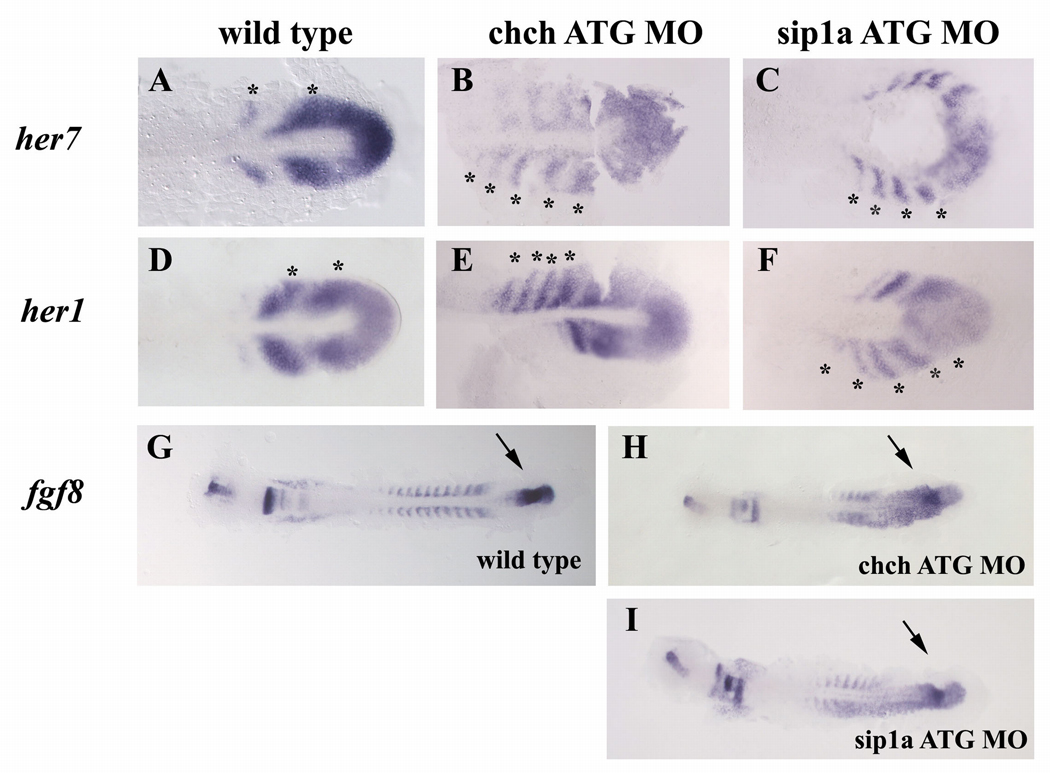
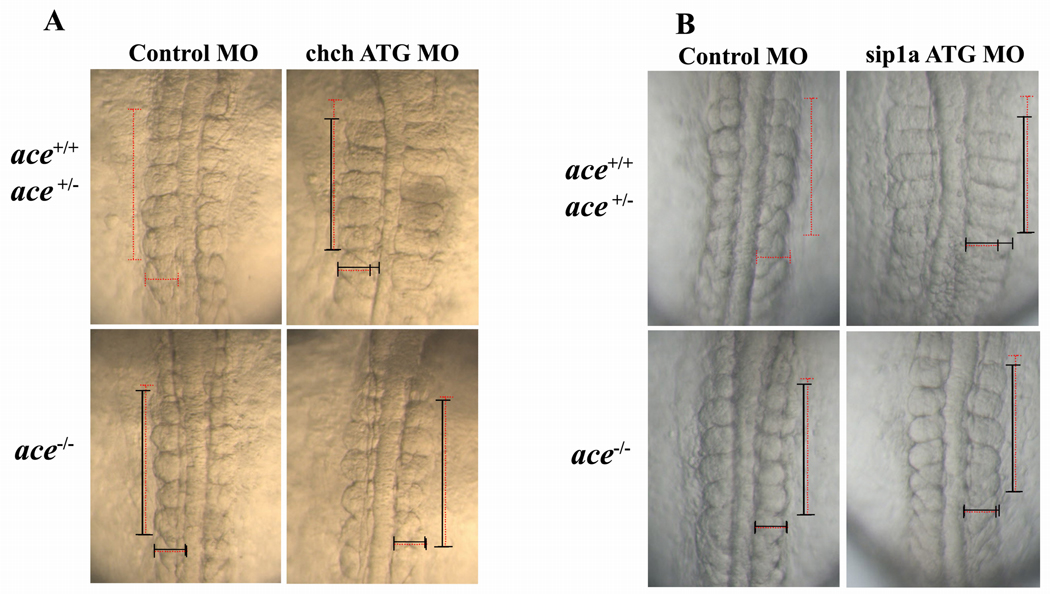
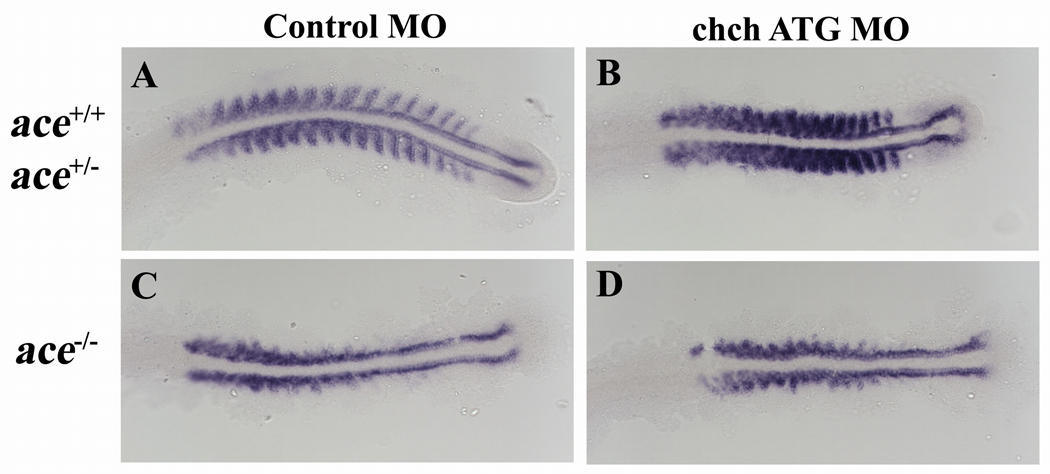
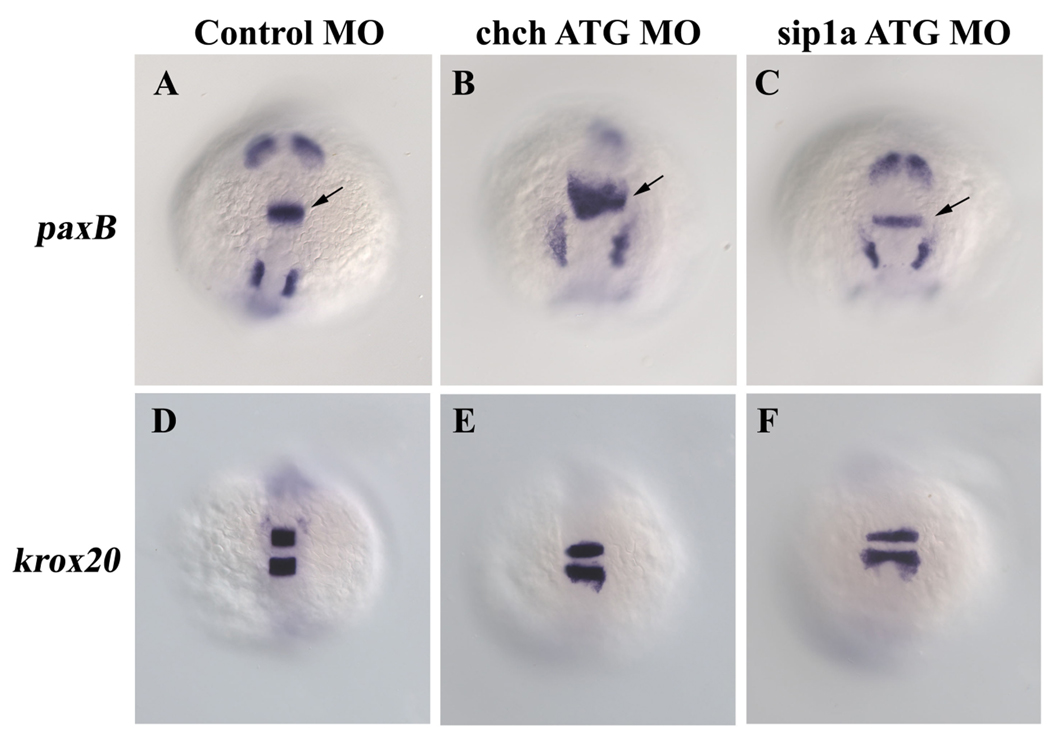
Cell-type specific regulation of a small number of growth factor signal transduction pathways generates diverse developmental outcomes. The zinc finger protein Churchill (ChCh) is a key effector of fibroblast growth factor (FGF) signaling during gastrulation. ChCh is largely thought to act by inducing expression of the multifunctional Sip1 (Smad Interacting Protein 1). We investigated the function of ChCh and Sip1a during zebrafish somitogenesis. Knockdown of ChCh or Sip1a results in misshapen somites that are short and narrow. As in wild-type embryos, cycling gene expression occurs in the developing somites in ChCh and Sip1a compromised embryos, but expression of her1 and her7 is maintained in formed somites. In addition, tail bud fgf8 expression is expanded anteriorly in these embryos. Finally, we found that blocking FGF8 restores somite morphology in ChCh and Sip1a compromised embryos. These results demonstrate a novel role for ChCh and Sip1a in repression of FGF activity.
Figures
References
-
- Amaya E, Musci TJ, Kirschner MW. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell. 1991;66:257–270. - PubMed
-
- Aulehla A, Wehrle C, Brand-Saberi B, Kemler R, Gossler A, Kanzler B, Herrmann BG. Wnt3a plays a major role in the segmentation clock controlling somitogenesis. Dev Cell. 2003;4:395–406. - PubMed
-
- Borland CZ, Schutzman JL, Stern MJ. Fibroblast growth factor signaling in Caenorhabditis elegans. Bioessays. 2001;23:1120–1130. - PubMed
-
- Bottcher RT, Niehrs C. Fibroblast growth factor signaling during early vertebrate development. Endocr Rev. 2005;26:63–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
