

Respiration in adipocytes is inhibited by reactive oxygen species
- PMID: 20035277
- PMCID: PMC6154476
- DOI: 10.1038/oby.2009.456
Respiration in adipocytes is inhibited by reactive oxygen species
Abstract
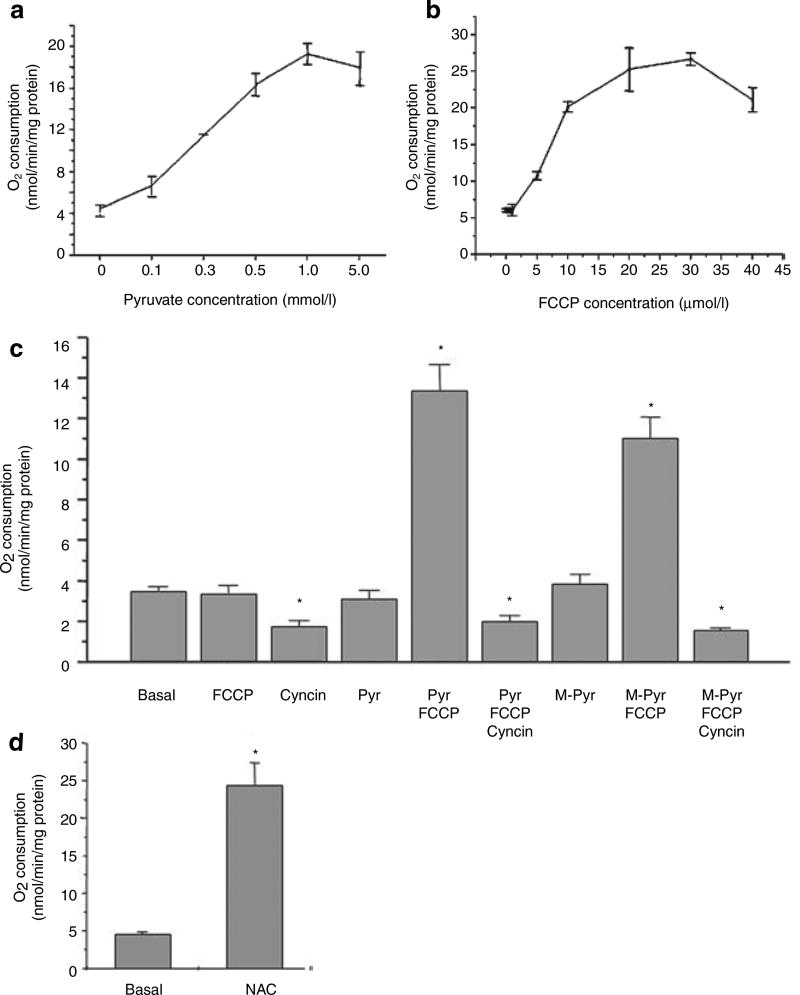
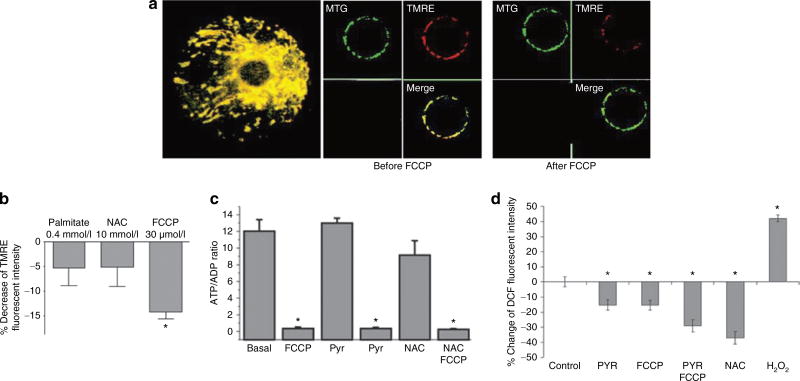
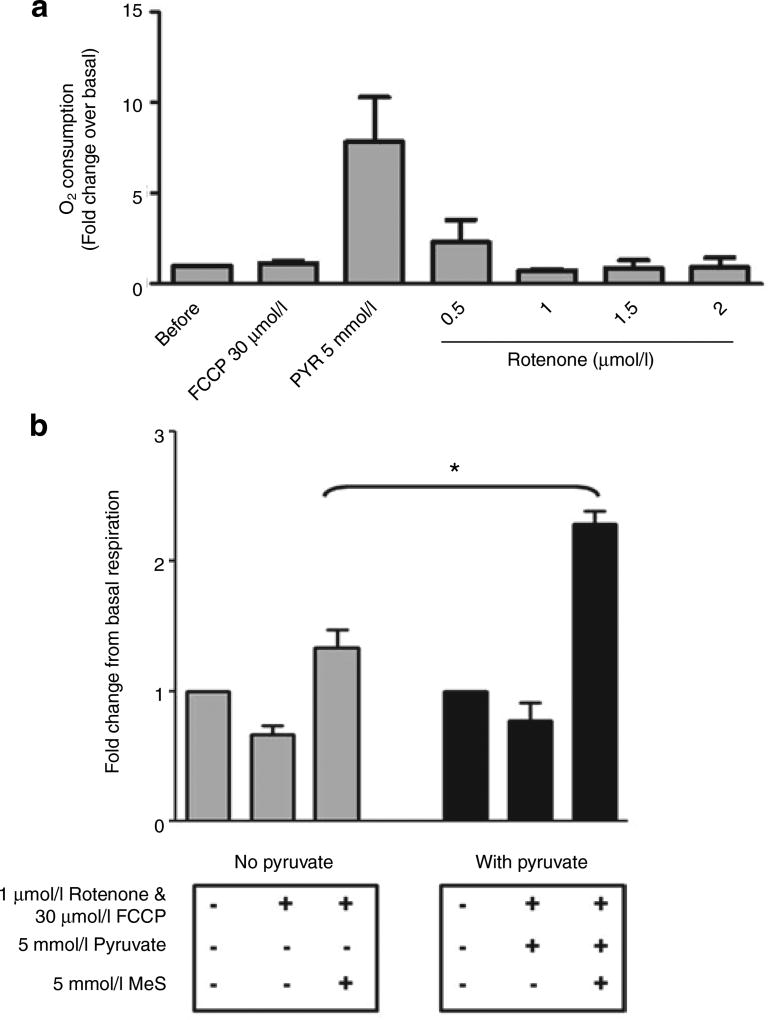
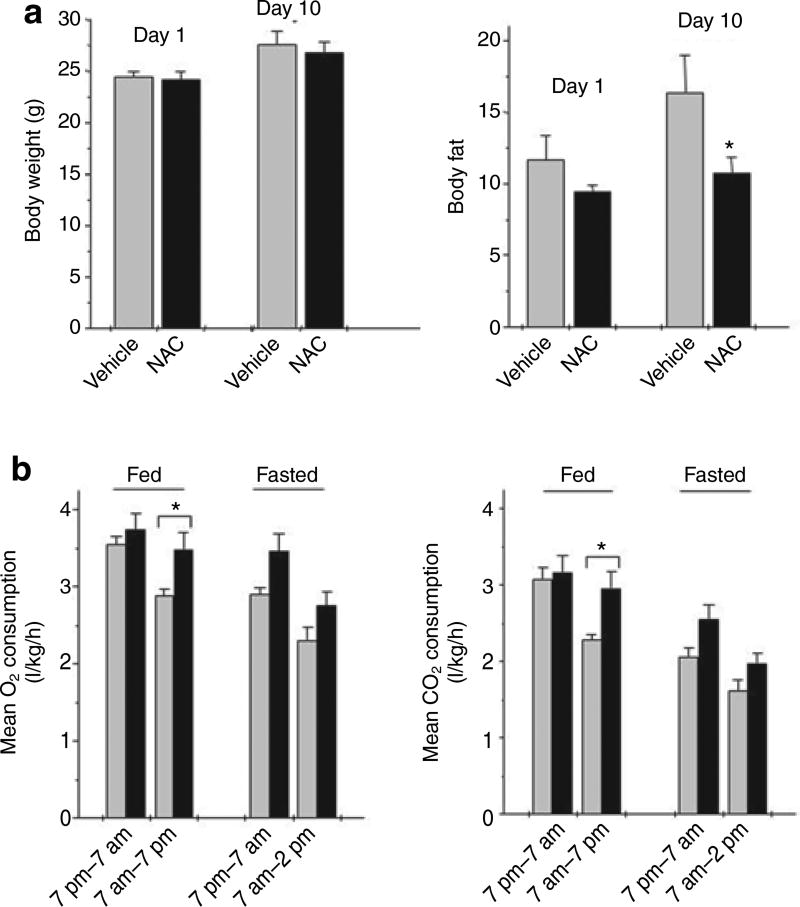
It is a desirable goal to stimulate fuel oxidation in adipocytes and shift the balance toward less fuel storage and more burning. To understand this regulatory process, respiration was measured in primary rat adipocytes, mitochondria, and fat-fed mice. Maximum O(2) consumption, in vitro, was determined with a chemical uncoupler of oxidative phosphorylation (carbonylcyanide p-trifluoromethoxyphenylhydrazone (FCCP)). The adenosine triphosphate/adenosine diphosphate (ATP/ADP) ratio was measured by luminescence. Mitochondria were localized by confocal microscopy with MitoTracker Green and their membrane potential (Delta psi(M)) measured using tetramethylrhodamine ethyl ester perchlorate (TMRE). The effect of N-acetylcysteine (NAC) on respiration and body composition in vivo was assessed in mice. Addition of FCCP collapsed Delta psi(M) and decreased the ATP/ADP ratio. However, we demonstrated the same rate of adipocyte O(2) consumption in the absence or presence of fuels and FCCP. Respiration was only stimulated when reactive oxygen species (ROS) were scavenged by pyruvate or NAC: other fuels or fuel combinations had little effect. Importantly, the ROS scavenging role of pyruvate was not affected by rotenone, an inhibitor of mitochondrial complex I. In addition, mice that consumed NAC exhibited increased O(2) consumption and decreased body fat in vivo. These studies suggest for the first time that adipocyte O(2) consumption may be inhibited by ROS, because pyruvate and NAC stimulated respiration. ROS inhibition of O(2) consumption may explain the difficulty to identify effective strategies to increase fat burning in adipocytes. Stimulating fuel oxidation in adipocytes by decreasing ROS may provide a novel means to shift the balance from fuel storage to fuel burning.
Conflict of interest statement
The authors declared no conflict of interest.
Figures
References
-
- McGarry JD, Takabayashi Y, Foster DW. The role of malonyl-coa in the coordination of fatty acid synthesis and oxidation in isolated rat hepatocytes. J Biol Chem. 1978;253:8294–8300. - PubMed
-
- Wang YL, Guo W, Zang Y, et al. Acyl coenzyme a synthetase regulation: putative role in long-chain acyl coenzyme a partitioning. Obes Res. 2004;12:1781–1788. - PubMed
-
- Wang T, Zang Y, Ling W, Corkey BE, Guo W. Metabolic partitioning of endogenous fatty acid in adipocytes. Obes Res. 2003;11:880–887. - PubMed
-
- Zang Y, Wang T, Xie W, et al. Regulation of acetyl CoA carboxylase and carnitine palmitoyl transferase-1 in rat adipocytes. Obes Res. 2005;13:1530–1539. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
