

Amyloid beta from axons and dendrites reduces local spine number and plasticity
- PMID: 20037574
- PMCID: PMC3310198
- DOI: 10.1038/nn.2476
Amyloid beta from axons and dendrites reduces local spine number and plasticity
Abstract
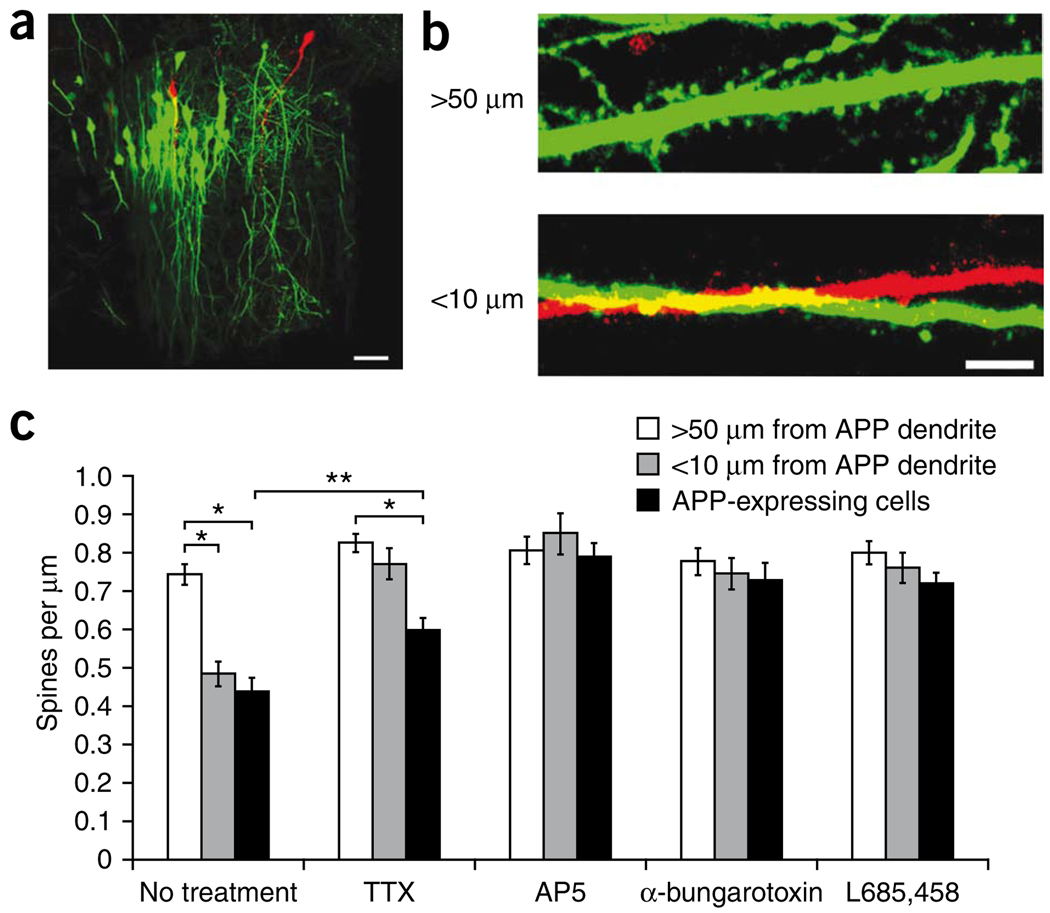
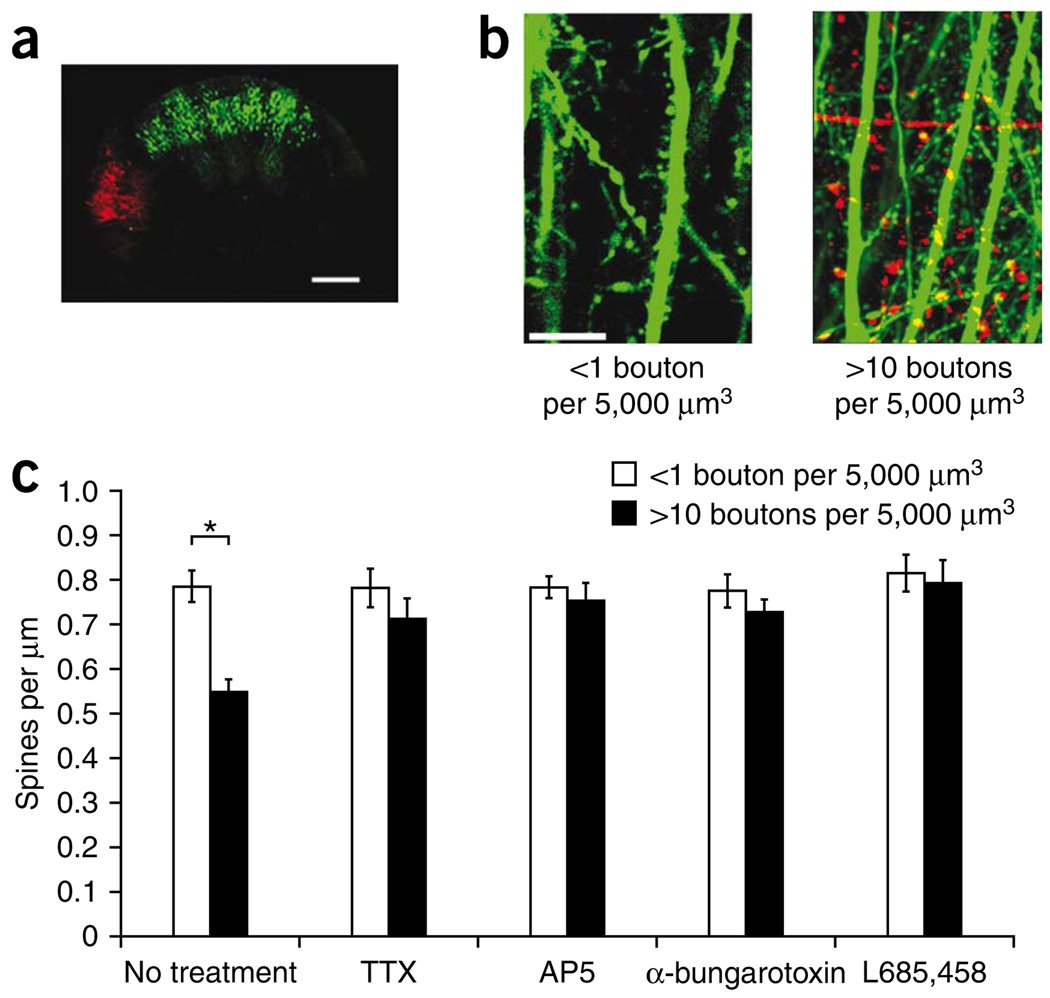
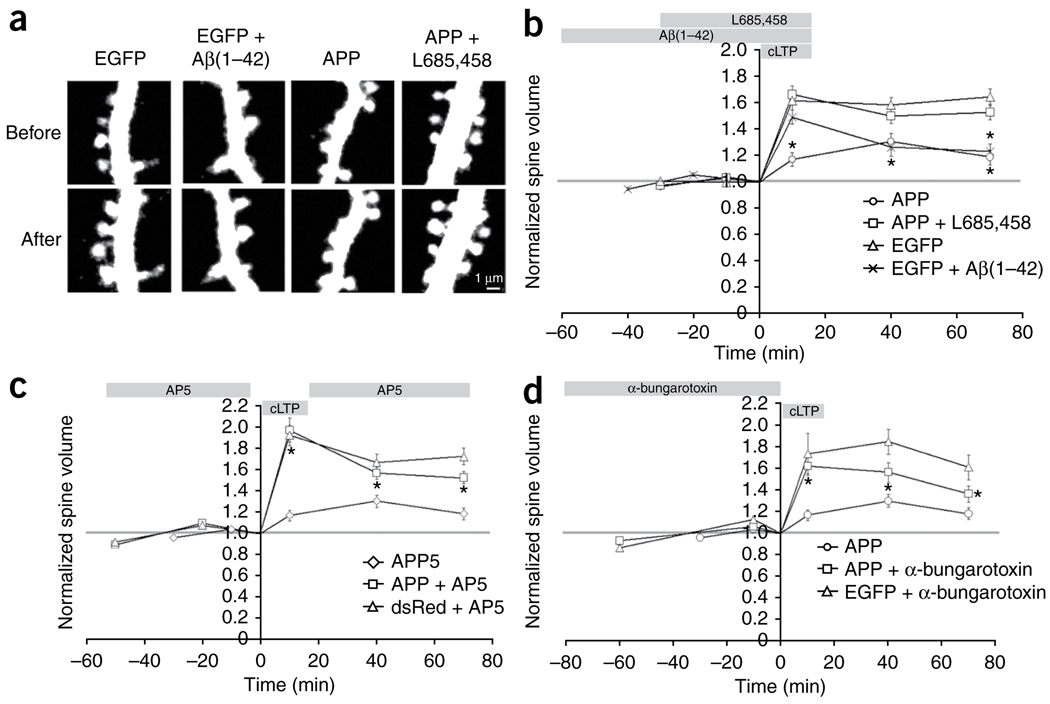
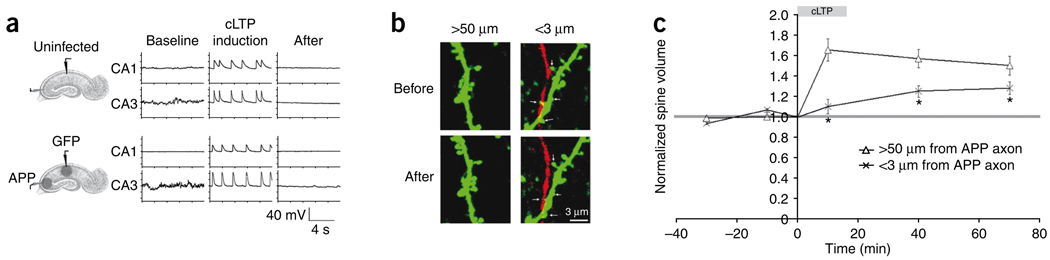
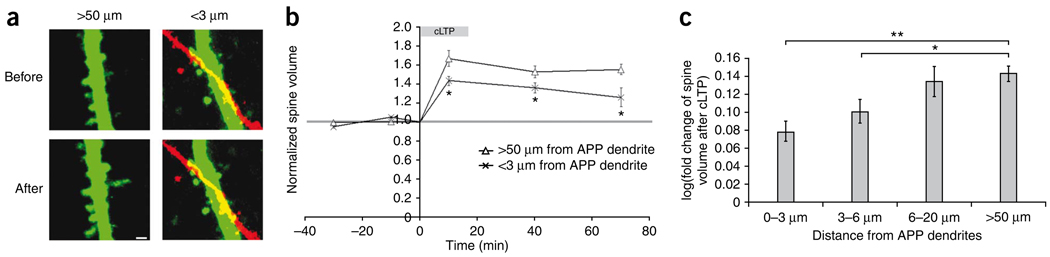
Excessive synaptic loss is thought to be one of the earliest events in Alzheimer's disease. Amyloid beta (Abeta), a peptide secreted in an activity-modulated manner by neurons, has been implicated in the pathogenesis of Alzheimer's disease by removing dendritic spines, sites of excitatory synaptic transmission. However, issues regarding the subcellular source of Abeta, as well as the mechanisms of its production and actions that lead to synaptic loss, remain poorly understood. In rat organotypic slices, we found that acute overproduction of either axonal or dendritic Abeta reduced spine density and plasticity at nearby ( approximately 5-10 mum) dendrites. The production of Abeta and its effects on spines were sensitive to blockade of action potentials or nicotinic receptors; the effects of Abeta (but not its production) were sensitive to NMDA receptor blockade. Notably, only 30-60 min blockade of Abeta overproduction permitted induction of plasticity. Our results indicate that continuous overproduction of Abeta at dendrites or axons acts locally to reduce the number and plasticity of synapses.
Figures


References
-
- Terry RD, et al. Physical basis of cognitive alterations in Alzheimer’s disease: synapse loss is the major correlate of cognitive impairment. Ann. Neurol. 1991;30:572–580. - PubMed
-
- Walsh DM, Selkoe DJ. Deciphering the molecular basis of memory failure in Alzheimer’s disease. Neuron. 2004;44:181–193. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
