

Isoform-specific contributions of alpha-actinin to glioma cell mechanobiology
- PMID: 20037648
- PMCID: PMC2793025
- DOI: 10.1371/journal.pone.0008427
Isoform-specific contributions of alpha-actinin to glioma cell mechanobiology
Abstract
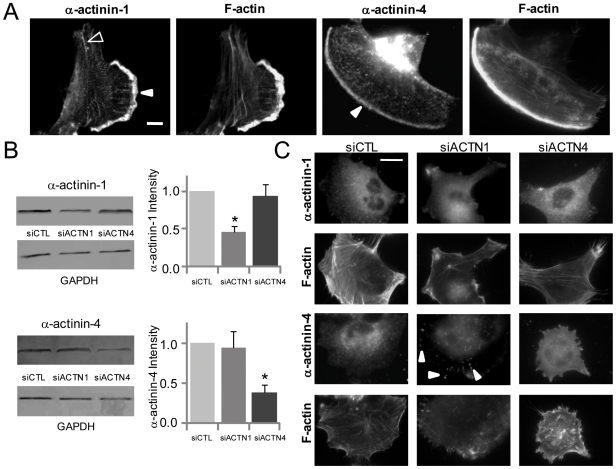
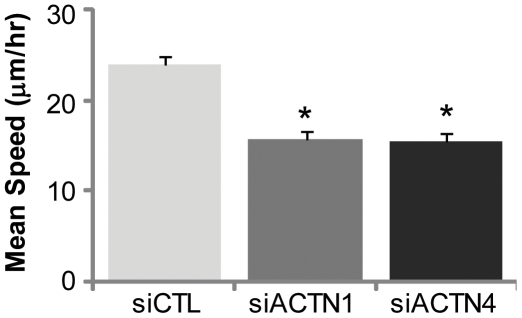
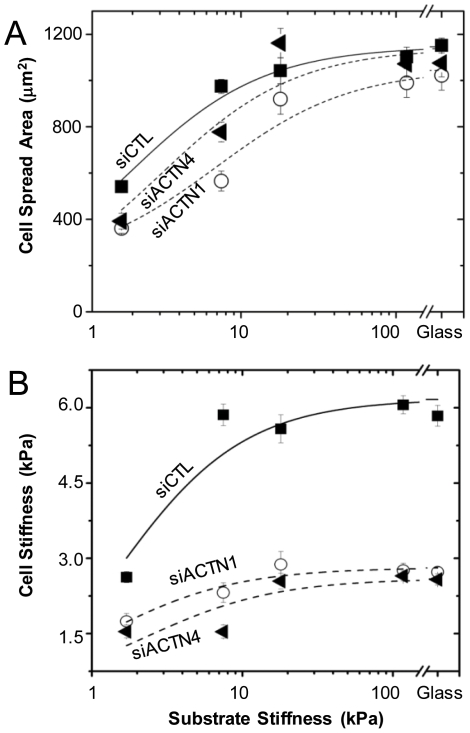
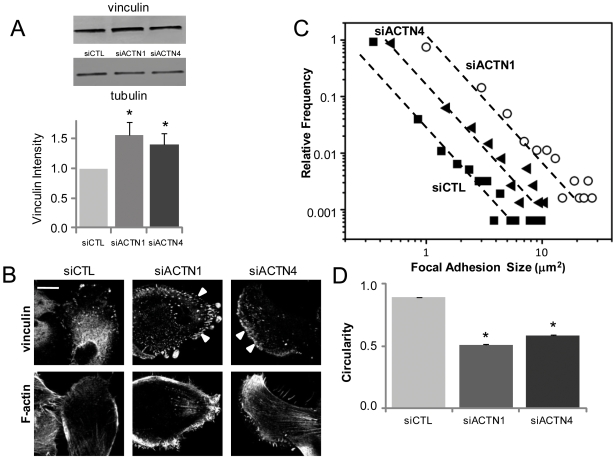
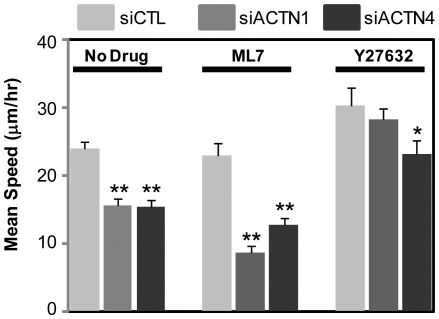
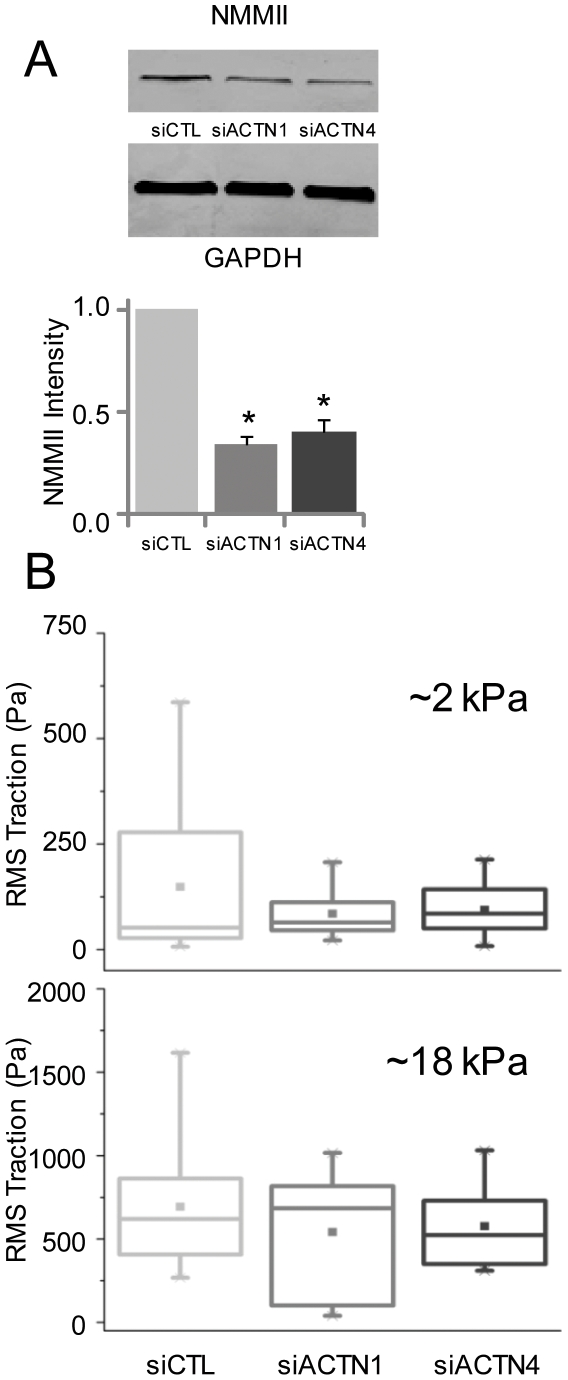
Glioblastoma Multiforme (GBM) is a malignant astrocytic tumor associated with low survival rates because of aggressive infiltration of tumor cells into the brain parenchyma. Expression of the actin binding protein alpha-actinin has been strongly correlated with the invasive phenotype of GBM in vivo. To probe the cellular basis of this correlation, we have suppressed expression of the nonmuscle isoforms alpha-actinin-1 and alpha-actinin-4 and examined the contribution of each isoform to the structure, mechanics, and motility of human glioma tumor cells in culture. While subcellular localization of each isoform is distinct, suppression of either isoform yields a phenotype that includes dramatically reduced motility, compensatory upregulation and redistribution of vinculin, reduced cortical elasticity, and reduced ability to adapt to changes in the elasticity of the extracellular matrix (ECM). Mechanistic studies reveal a relationship between alpha-actinin and non-muscle myosin II in which depletion of either alpha-actinin isoform reduces myosin expression and maximal cell-ECM tractional forces. Our results demonstrate that both alpha-actinin-1 and alpha-actinin-4 make critical and distinct contributions to cytoskeletal organization, rigidity-sensing, and motility of glioma cells, thereby yielding mechanistic insight into the observed correlation between alpha-actinin expression and GBM invasiveness in vivo.
Conflict of interest statement
Figures
Similar articles
-
The microtubule inhibiting agent epothilone B antagonizes glioma cell motility associated with reorganization of the actin-binding protein α-actinin 4.Oncol Rep. 2011 Mar;25(3):887-93. doi: 10.3892/or.2011.1145. Epub 2011 Jan 13. Oncol Rep. 2011. PMID: 21234524
-
The mechanical rigidity of the extracellular matrix regulates the structure, motility, and proliferation of glioma cells.Cancer Res. 2009 May 15;69(10):4167-74. doi: 10.1158/0008-5472.CAN-08-4859. Epub 2009 May 12. Cancer Res. 2009. PMID: 19435897 Free PMC article.
-
Molecular characterization of cell substratum attachments in human glial tumors relates to prognostic features.Glia. 2001 Dec;36(3):375-90. doi: 10.1002/glia.1124. Glia. 2001. PMID: 11746774
-
Microscopic co-distributions of myosin, actin, alpha-actinin and vinculin in human neutrophils during movement.Haematologia (Budap). 1999;29(4):285-94. Haematologia (Budap). 1999. PMID: 10438068
-
Mechanotransduction pathways linking the extracellular matrix to the nucleus.Int Rev Cell Mol Biol. 2014;310:171-220. doi: 10.1016/B978-0-12-800180-6.00005-0. Int Rev Cell Mol Biol. 2014. PMID: 24725427 Review.
Cited by
-
Strategies of eradicating glioma cells: a multi-scale mathematical model with MiR-451-AMPK-mTOR control.PLoS One. 2015 Jan 28;10(1):e0114370. doi: 10.1371/journal.pone.0114370. eCollection 2015. PLoS One. 2015. PMID: 25629604 Free PMC article.
-
Engineering strategies to mimic the glioblastoma microenvironment.Adv Drug Deliv Rev. 2014 Dec 15;79-80:172-83. doi: 10.1016/j.addr.2014.08.012. Epub 2014 Aug 29. Adv Drug Deliv Rev. 2014. PMID: 25174308 Free PMC article. Review.
-
Influence of Inherent Mechanophenotype on Competitive Cellular Adherence.Ann Biomed Eng. 2017 Aug;45(8):2036-2047. doi: 10.1007/s10439-017-1841-5. Epub 2017 Apr 26. Ann Biomed Eng. 2017. PMID: 28447179 Free PMC article.
-
Role of Microenvironment in Glioma Invasion: What We Learned from In Vitro Models.Int J Mol Sci. 2018 Jan 4;19(1):147. doi: 10.3390/ijms19010147. Int J Mol Sci. 2018. PMID: 29300332 Free PMC article. Review.
-
The role of myosin II in glioma invasion: A mathematical model.PLoS One. 2017 Feb 6;12(2):e0171312. doi: 10.1371/journal.pone.0171312. eCollection 2017. PLoS One. 2017. PMID: 28166231 Free PMC article.
References
-
- Furnari FB, Fenton T, Bachoo RM, Mukasa A, Stommel JM, et al. Malignant astrocytic glioma: genetics, biology, and paths to treatment. Genes & Development. 2007;21:2683–2710. - PubMed
-
- Fukushima Y, Ohnishi T, Arita N, Hayakawa T, Sekiguchi K. Integrin alpha3beta1-mediated interaction with laminin-5 stimulates adhesion, migration and invasion of malignant glioma cells. Int J Cancer. 1998;76:63–72. - PubMed
-
- Rao JS. Molecular mechanisms of glioma invasiveness: The role of proteases. Nature Reviews Cancer. 2003;3:489–501. - PubMed
-
- Demuth T, Berens ME. Molecular mechanisms of glioma cell migration and invasion. Journal of Neuro-Oncology. 2004;70:217–228. - PubMed
-
- Wolf K, Wu YI, Liu Y, Geiger J, Tam E, et al. Multi-step pericellular proteolysis controls the transition from individual to collective cancer cell invasion. Nat Cell Biol. 2007;9:893–904. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
