

Rates and mechanisms of resistance development in Mycobacterium tuberculosis to a novel diarylquinoline ATP synthase inhibitor
- PMID: 20038615
- PMCID: PMC2825986
- DOI: 10.1128/AAC.01611-09
Rates and mechanisms of resistance development in Mycobacterium tuberculosis to a novel diarylquinoline ATP synthase inhibitor
Abstract
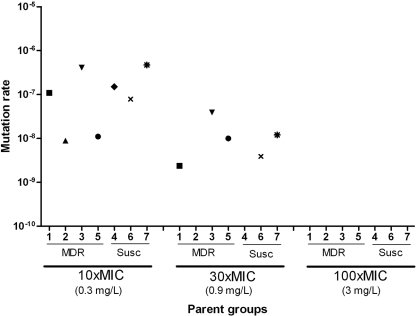
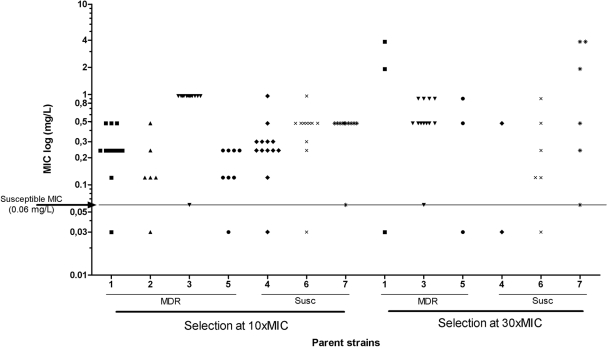
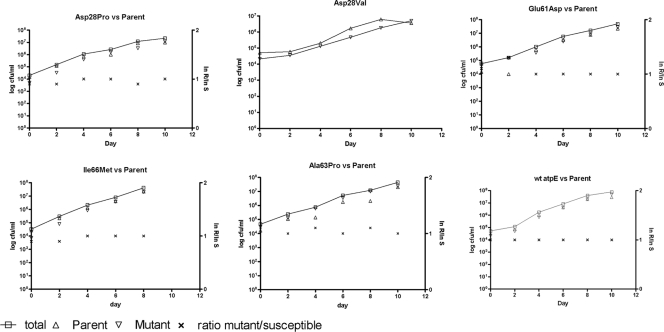
R207910 (also known as TMC207) is an investigational drug currently in clinical studies for the treatment of multidrug-resistant (MDR) tuberculosis. It has a high degree of antimycobacterial activity and is equally effective against drug-susceptible and MDR Mycobacterium tuberculosis isolates. In the present study, we characterized the development of resistance to R207910 in vitro. Ninety-seven independent R207910-resistant mutants were selected from seven different clinical isolates of M. tuberculosis (three drug-susceptible and four MDR isolates) at 10x, 30x, and 100x the MIC. At a concentration of 0.3 mg/liter (10x the MIC), the mutation rates ranged from 4.7 x 10(-7) to 8.9 x 10(-9) mutations per cell per division, and at 1.0 mg/liter (30x the MIC) the mutation rate ranged from 3.9 x 10(-8) to 2.4 x 10(-9). No resistant mutants were obtained at 3 mg/liter (100x the MIC). The level of resistance ranged from 0.12 to 3.84 mg/liter for the mutants identified; these concentrations represent 4- to 128-fold increases in the MICs. For 53 of the resistant mutants, the atpE gene, which encodes a transmembrane and oligomeric C subunit of the ATP synthase and which was previously shown to be involved in resistance, was sequenced. For 15/53 mutants, five different point mutations resulting in five different amino acid substitutions were identified in the atpE gene. For 38/53 mutants, no atpE mutations were found and sequencing of the complete F0 ATP synthase operon (atpB, atpE, and atpF genes) and the F1 ATP synthase operon (atpH, atpA, atpG, atpD, and atpC genes) from three mutants revealed no mutations, indicating other, alternative resistance mechanisms. Competition assays showed no measurable reduction in the fitness of the mutants compared to that of the isogenic wild types.
Figures
References
-
- Andersson, D. I. 2006. The biological cost of mutational antibiotic resistance: any practical conclusions? Curr. Opin. Microbiol. 9:461-465. - PubMed
-
- Andersson, D. I. 2003. Persistence of antibiotic resistant bacteria. Curr. Opin. Microbiol. 6:452-456. - PubMed
-
- Andersson, D. I., and B. R. Levin. 1999. The biological cost of antibiotic resistance. Curr. Opin. Microbiol. 2:489-493. - PubMed
-
- Andries, K., P. Verhasselt, J. Guillemont, H. W. Gohlmann, J. M. Neefs, H. Winkler, J. Van Gestel, P. Timmerman, M. Zhu, E. Lee, P. Williams, D. de Chaffoy, E. Huitric, S. Hoffner, E. Cambau, C. Truffot-Pernot, N. Lounis, and V. Jarlier. 2005. A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. Science 307:223-227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
