

Genetic and epigenetic silencing of SCARA5 may contribute to human hepatocellular carcinoma by activating FAK signaling
- PMID: 20038795
- PMCID: PMC2798676
- DOI: 10.1172/JCI38012
Genetic and epigenetic silencing of SCARA5 may contribute to human hepatocellular carcinoma by activating FAK signaling
Abstract
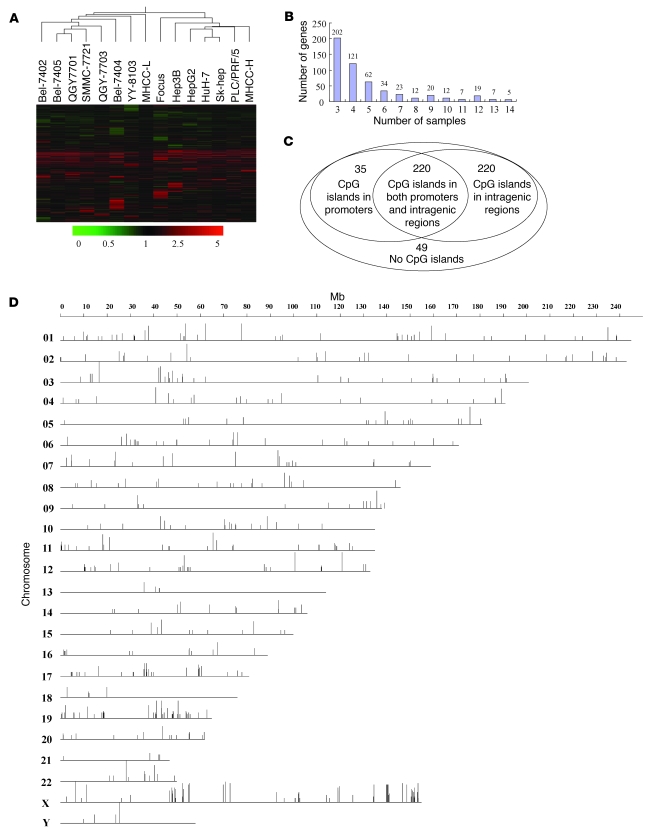
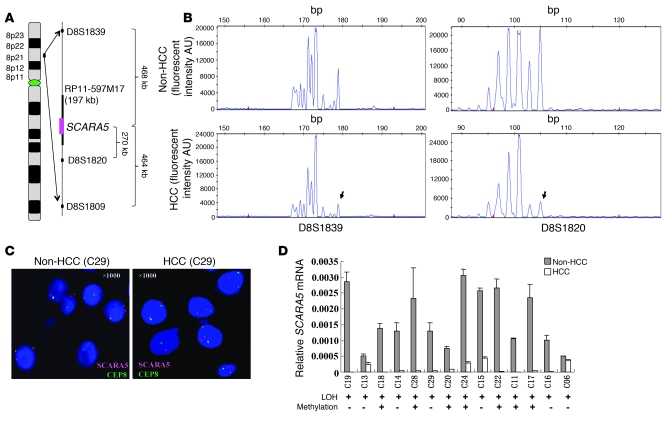
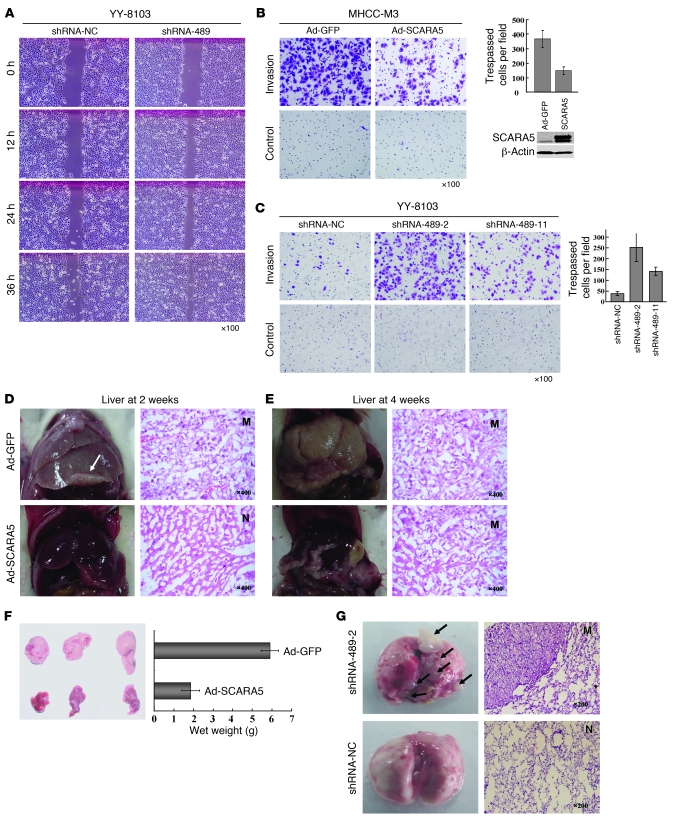
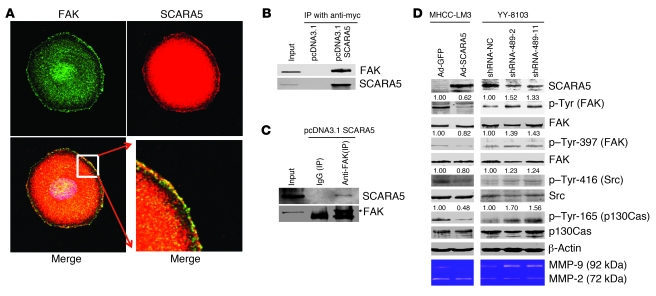
The epigenetic silencing of tumor suppressor genes is a crucial event during carcinogenesis and metastasis. Here, in a human genome-wide survey, we identified scavenger receptor class A, member 5 (SCARA5) as a candidate tumor suppressor gene located on chromosome 8p. We found that SCARA5 expression was frequently downregulated as a result of promoter hypermethylation and allelic imbalance and was associated with vascular invasion in human hepatocellular carcinoma (HCC). Furthermore, SCARA5 knockdown via RNAi markedly enhanced HCC cell growth in vitro, colony formation in soft agar, and invasiveness, tumorigenicity, and lung metastasis in vivo. By contrast, SCARA5 overexpression suppressed these malignant behaviors. Interestingly, SCARA5 was found to physically associate with focal adhesion kinase (FAK) and inhibit the tyrosine phosphorylation cascade of the FAK-Src-Cas signaling pathway. Conversely, silencing SCARA5 stimulated the signaling pathway via increased phosphorylation of certain tyrosine residues of FAK, Src, and p130Cas; it was also associated with activation of MMP9, a tumor metastasis-associated enzyme. Taken together, these data suggest that the plasma membrane protein SCARA5 can contribute to HCC tumorigenesis and metastasis via activation of the FAK signaling pathway.
Figures






Similar articles
-
SPAG5 promotes hepatocellular carcinoma progression by downregulating SCARA5 through modifying β-catenin degradation.J Exp Clin Cancer Res. 2018 Sep 18;37(1):229. doi: 10.1186/s13046-018-0891-3. J Exp Clin Cancer Res. 2018. PMID: 30249289 Free PMC article.
-
Therapeutic upregulation of Class A scavenger receptor member 5 inhibits tumor growth and metastasis.Cancer Sci. 2012 Sep;103(9):1631-9. doi: 10.1111/j.1349-7006.2012.02350.x. Epub 2012 Aug 17. Cancer Sci. 2012. PMID: 22642751 Free PMC article.
-
Overexpression of SCARA5 inhibits tumor proliferation and invasion in osteosarcoma via suppression of the FAK signaling pathway.Mol Med Rep. 2016 Mar;13(3):2885-91. doi: 10.3892/mmr.2016.4857. Epub 2016 Feb 3. Mol Med Rep. 2016. PMID: 26847210
-
Capn4 contributes to tumour growth and metastasis of hepatocellular carcinoma by activation of the FAK-Src signalling pathways.J Pathol. 2014 Nov;234(3):316-28. doi: 10.1002/path.4395. Epub 2014 Aug 18. J Pathol. 2014. PMID: 24962955
-
The role of focal adhesion kinase in tumor initiation and progression.Cell Adh Migr. 2009 Oct-Dec;3(4):347-50. doi: 10.4161/cam.3.4.9458. Epub 2009 Oct 3. Cell Adh Migr. 2009. PMID: 19690467 Free PMC article. Review.
Cited by
-
G9a/DNMT1 co-targeting inhibits non-small cell lung cancer growth and reprograms tumor cells to respond to cancer-drugs through SCARA5 and AOX1.Cell Death Dis. 2024 Nov 2;15(11):787. doi: 10.1038/s41419-024-07156-w. Cell Death Dis. 2024. PMID: 39488528 Free PMC article.
-
Genomic alterations on 8p21-p23 are the most frequent genetic events in stage I squamous cell carcinoma of the lung.Exp Ther Med. 2015 Feb;9(2):345-350. doi: 10.3892/etm.2014.2123. Epub 2014 Dec 9. Exp Ther Med. 2015. PMID: 25574196 Free PMC article.
-
SCARA3 inhibits cell proliferation and EMT through AKT signaling pathway in lung cancer.BMC Cancer. 2022 May 16;22(1):552. doi: 10.1186/s12885-022-09631-z. BMC Cancer. 2022. PMID: 35578316 Free PMC article.
-
SPAG5 promotes hepatocellular carcinoma progression by downregulating SCARA5 through modifying β-catenin degradation.J Exp Clin Cancer Res. 2018 Sep 18;37(1):229. doi: 10.1186/s13046-018-0891-3. J Exp Clin Cancer Res. 2018. PMID: 30249289 Free PMC article.
-
PepFect14 Signaling and Transfection.Methods Mol Biol. 2022;2383:229-246. doi: 10.1007/978-1-0716-1752-6_15. Methods Mol Biol. 2022. PMID: 34766293
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
