

Limited forward trafficking of connexin 43 reduces cell-cell coupling in stressed human and mouse myocardium
- PMID: 20038810
- PMCID: PMC2798685
- DOI: 10.1172/JCI39740
Limited forward trafficking of connexin 43 reduces cell-cell coupling in stressed human and mouse myocardium
Abstract
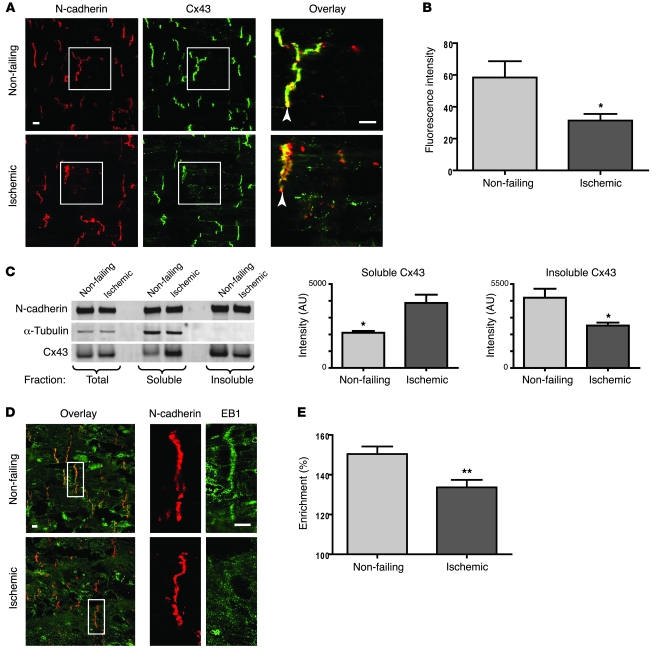
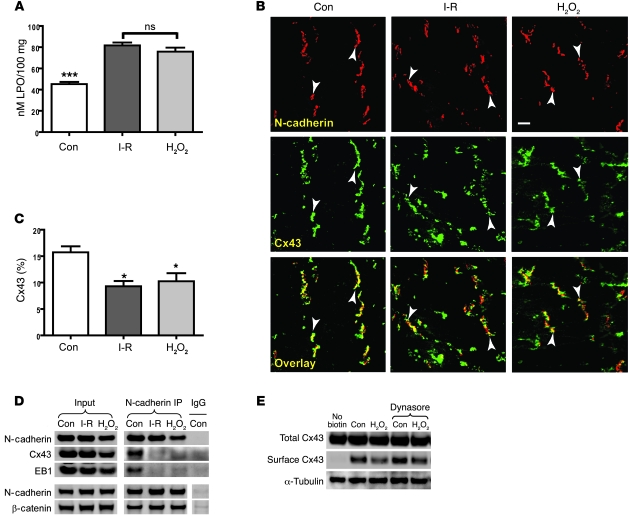
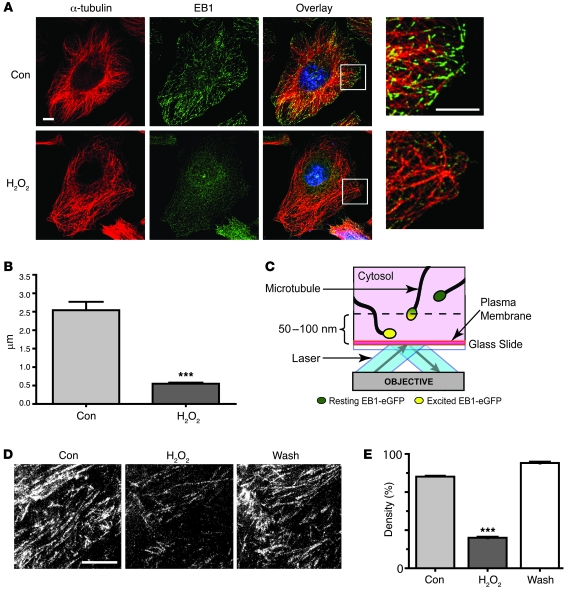
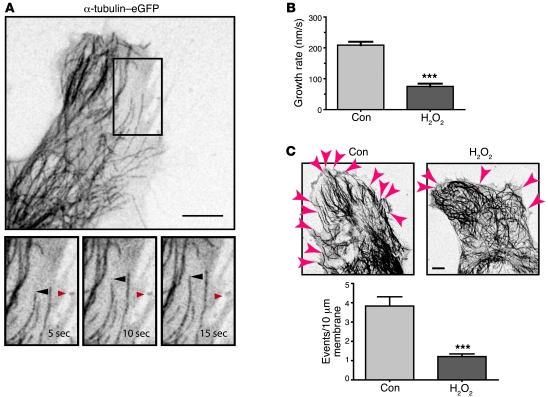
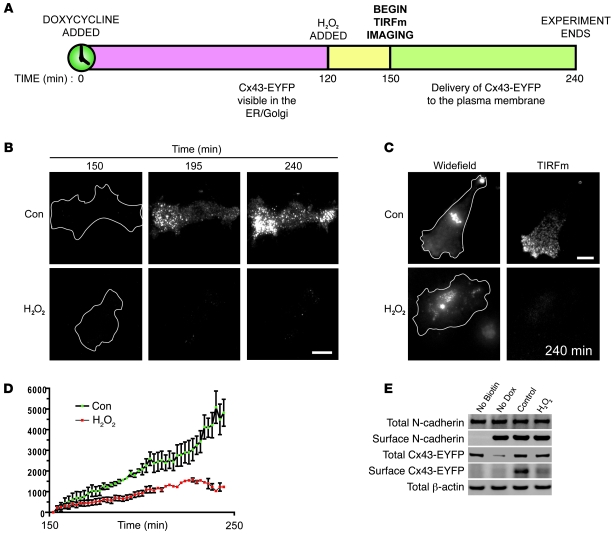
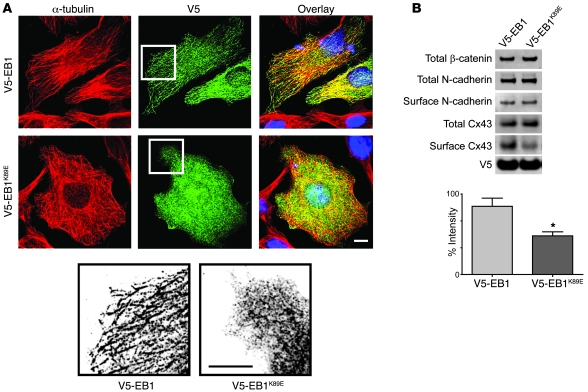
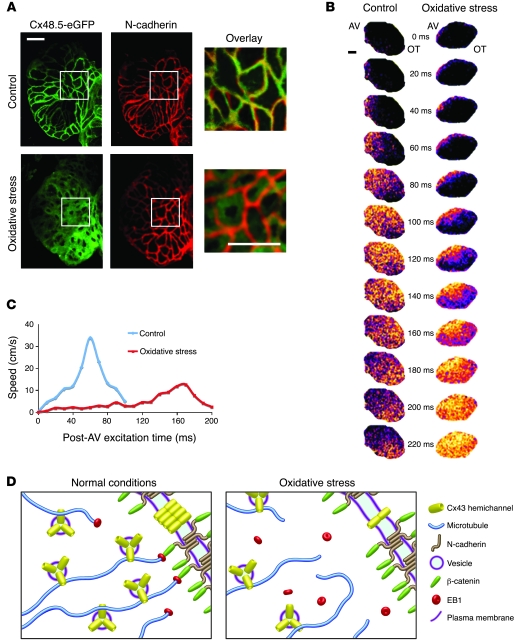
Gap junctions form electrical conduits between adjacent myocardial cells, permitting rapid spatial passage of the excitation current essential to each heartbeat. Arrhythmogenic decreases in gap junction coupling are a characteristic of stressed, failing, and aging myocardium, but the mechanisms of decreased coupling are poorly understood. We previously found that microtubules bearing gap junction hemichannels (connexons) can deliver their cargo directly to adherens junctions. The specificity of this delivery requires the microtubule plus-end tracking protein EB1. We performed this study to investigate the hypothesis that the oxidative stress that accompanies acute and chronic ischemic disease perturbs connexon forward trafficking. We found that EB1 was displaced in ischemic human hearts, stressed mouse hearts, and isolated cells subjected to oxidative stress. As a result, we observed limited microtubule interaction with adherens junctions at intercalated discs and reduced connexon delivery and gap junction coupling. A point mutation within the tubulin-binding domain of EB1 reproduced EB1 displacement and diminished connexon delivery, confirming that EB1 displacement can limit gap junction coupling. In zebrafish hearts, oxidative stress also reduced the membrane localization of connexin and slowed the spatial spread of excitation. We anticipate that protecting the microtubule-based forward delivery apparatus of connexons could improve cell-cell coupling and reduce ischemia-related cardiac arrhythmias.
Figures
Comment in
-
Oxidant stress derails the cardiac connexon connection.J Clin Invest. 2010 Jan;120(1):87-9. doi: 10.1172/JCI41780. Epub 2009 Dec 28. J Clin Invest. 2010. PMID: 20038808 Free PMC article.
References
-
- Shaw RM, Rudy Y. Ionic mechanisms of propagation in cardiac tissue. Roles of the sodium and L-type calcium currents during reduced excitability and decreased gap junction coupling. Circ Res. 1997;81(5):727–741. - PubMed
-
- Beardslee MA, et al. Dephosphorylation and intracellular redistribution of ventricular connexin43 during electrical uncoupling induced by ischemia. Circ Res. 2000;87(8):656–662. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
