

Deep evolutionary roots of strepsirrhine primate labyrinthine morphology
- PMID: 20039977
- PMCID: PMC2829395
- DOI: 10.1111/j.1469-7580.2009.01177.x
Deep evolutionary roots of strepsirrhine primate labyrinthine morphology
Abstract
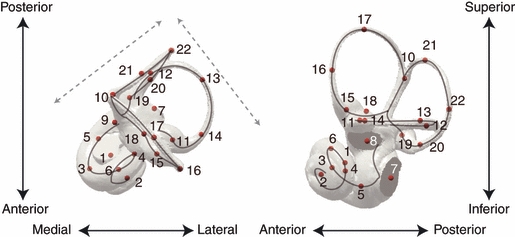
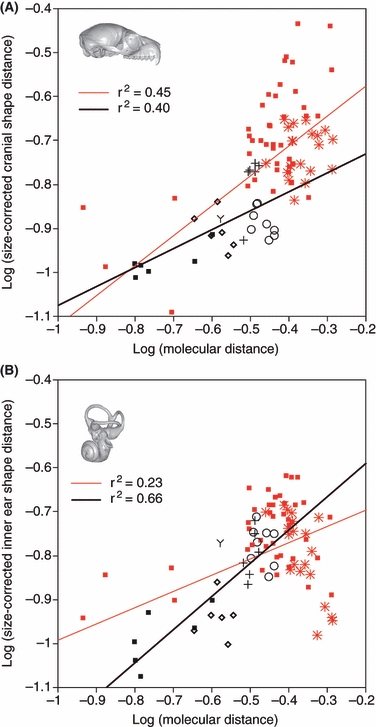
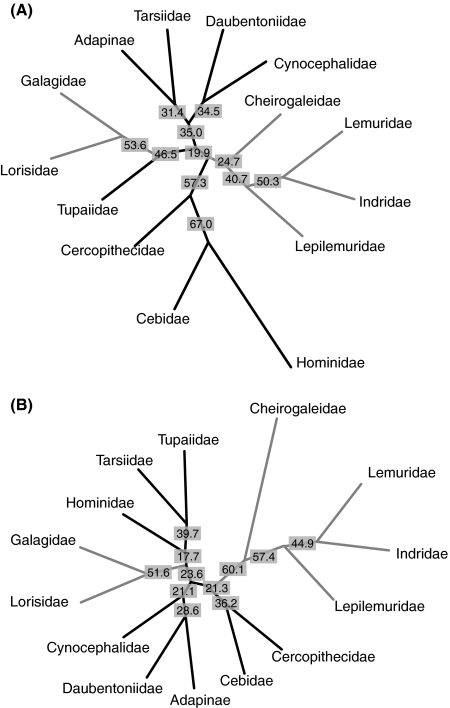
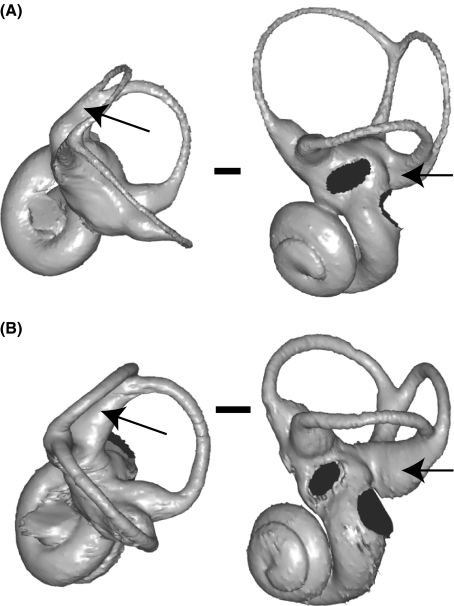
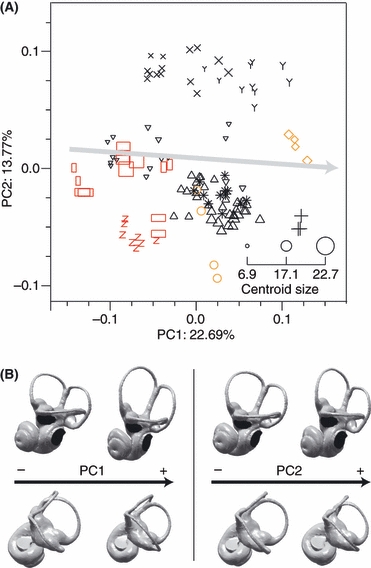
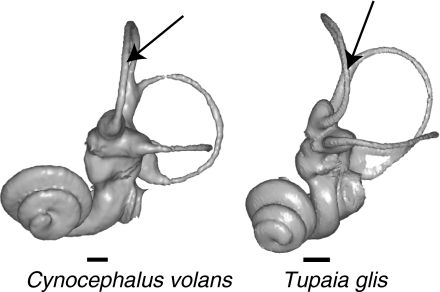
The cavity system of the inner ear of mammals is a complex three-dimensional structure that houses the organs of equilibrium and hearing. Morphological variation of the inner ear across mammals reflects differences in locomotor behaviour and hearing performance, and the good preservation of this structure in many fossil specimens permits analogous inferences. However, it is less well known to what extent the morphology of the bony labyrinth conveys information about the evolutionary history of primate taxa. We studied this question in strepsirrhine primates with the aim to assess the potential and limitations of using the inner ear as a phylogenetic marker. Geometric morphometric analysis showed that the labyrinthine morphology of extant strepsirrhines contains a mixed locomotor, allometric and phylogenetic signal. Discriminant analysis at the family level confirmed that labyrinthine shape is a good taxonomic marker. Our results support the hypothesis that evolutionary change in labyrinthine morphology is adequately described with a random walk model, i.e. random phenotypic dispersal in morphospace. Under this hypothesis, average shapes calculated for each node of the phylogenetic tree give an estimate of inner ear shapes of the respective last common ancestors (LCAs), and this information can be used to infer character state polarity. The labyrinthine morphology of the fossil Adapinae is close to the inferred basal morphology of the strepsirrhines. The inner ear of Daubentonia, one of the most derived extant strepsirrhines, is autapomorphic in many respects, but also presents unique similarities with adapine labyrinths.
Figures
References
-
- Amenta N, Choi S, Kolluri R. The Power Crust. Ann Arbor: Association for Computing Machinery; 2001. pp. 249–260. Proceedings of the Sixth ACM Symposium on Solid Modeling and Applications.
-
- Bacon A-M, Godinot M. Analyse morphofonctionnelle des fémurs et des tibias des ‘Adapis’ du Quercy: mise en évidence de cinq types morphologiques. Folia Primatol. 1998;69:1–21. - PubMed
-
- Charles-Dominique P, Martin RD. Evolution of lorises and lemurs. Nature. 1970;227:257–260. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
