

Tuning microbial hosts for membrane protein production
- PMID: 20040113
- PMCID: PMC2807855
- DOI: 10.1186/1475-2859-8-69
Tuning microbial hosts for membrane protein production
Abstract
The last four years have brought exciting progress in membrane protein research. Finally those many efforts that have been put into expression of eukaryotic membrane proteins are coming to fruition and enable to solve an ever-growing number of high resolution structures. In the past, many skilful optimization steps were required to achieve sufficient expression of functional membrane proteins. Optimization was performed individually for every membrane protein, but provided insight about commonly encountered bottlenecks and, more importantly, general guidelines how to alleviate cellular limitations during microbial membrane protein expression. Lately, system-wide analyses are emerging as powerful means to decipher cellular bottlenecks during heterologous protein production and their use in microbial membrane protein expression has grown in popularity during the past months.This review covers the most prominent solutions and pitfalls in expression of eukaryotic membrane proteins using microbial hosts (prokaryotes, yeasts), highlights skilful applications of our basic understanding to improve membrane protein production. Omics technologies provide new concepts to engineer microbial hosts for membrane protein production.
Figures
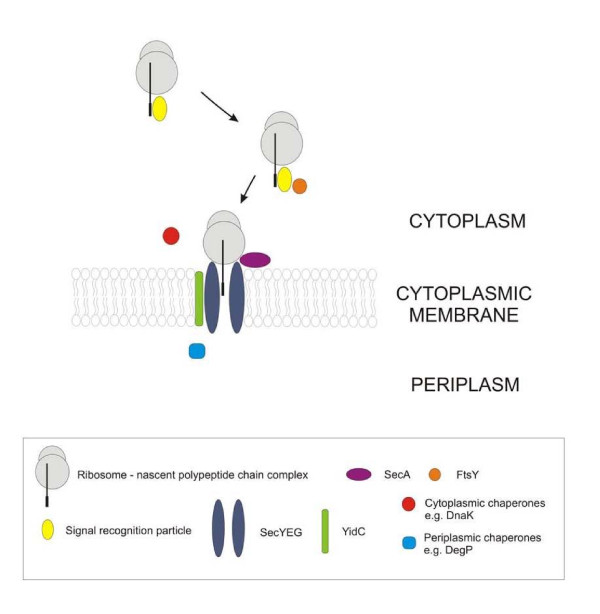
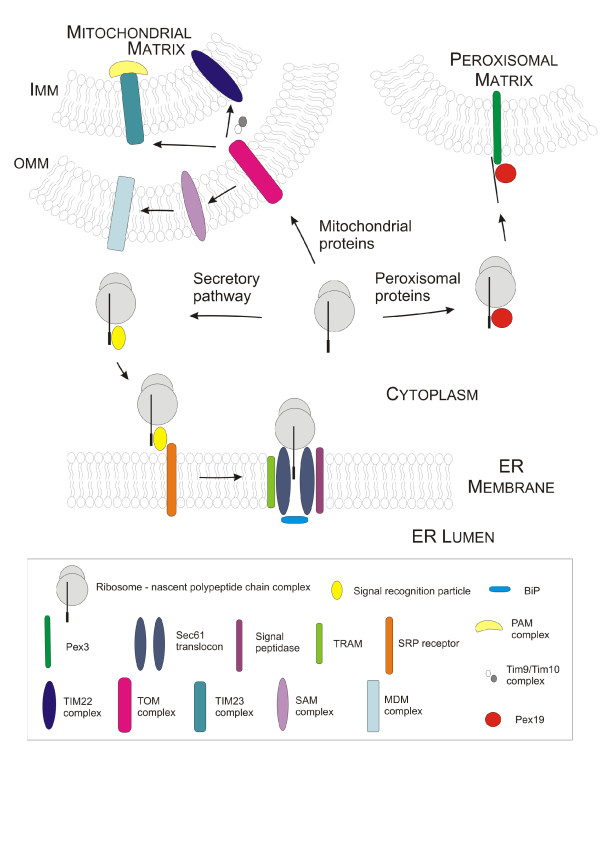
References
-
- Rasmussen SG, Choi HJ, Rosenbaum DM, Kobilka TS, Thian FS, Edwards PC, Burghammer M, Ratnala VR, Sanishvili R, Fischetti RF, Schertler GF, Weis WI, Kobilka BK. Crystal structure of the human beta2 adrenergic G-protein-coupled receptor. Nature. 2007;450:383–387. doi: 10.1038/nature06325. - DOI - PubMed
-
- Cherezov V, Rosenbaum DM, Hanson MA, Rasmussen SG, Thian FS, Kobilka TS, Choi HJ, Kuhn P, Weis WI, Kobilka BK, Stevens RC. High-resolution crystal structure of an engineered human beta2-adrenergic G protein-coupled receptor. Science. 2007;318:1258–1265. doi: 10.1126/science.1150577. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
