

Rearrangement of the RNA polymerase subunit H and the lower jaw in archaeal elongation complexes
- PMID: 20040576
- PMCID: PMC2847245
- DOI: 10.1093/nar/gkp1190
Rearrangement of the RNA polymerase subunit H and the lower jaw in archaeal elongation complexes
Abstract
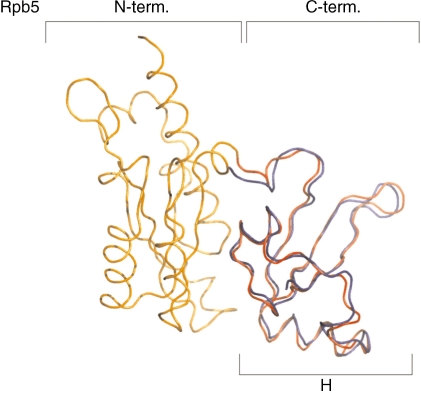
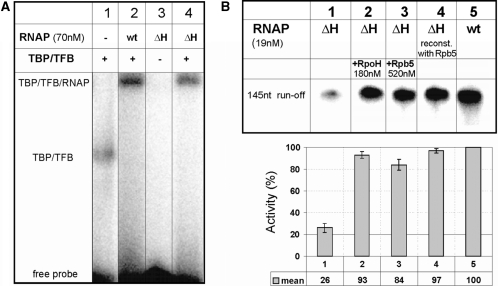
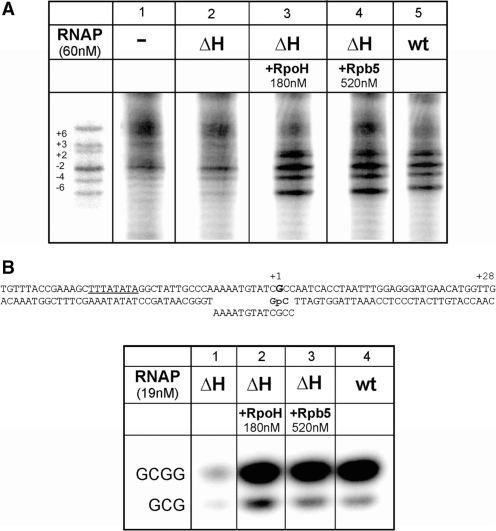
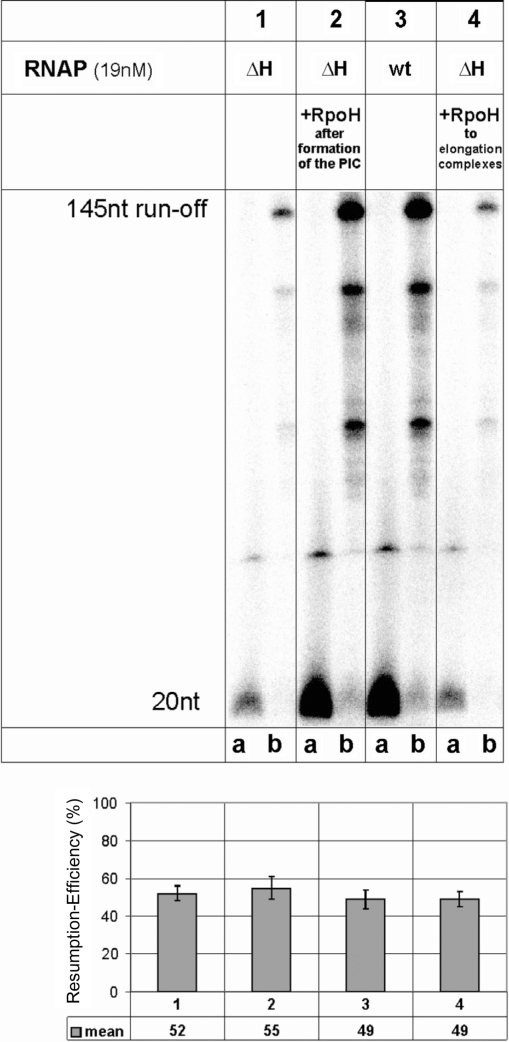
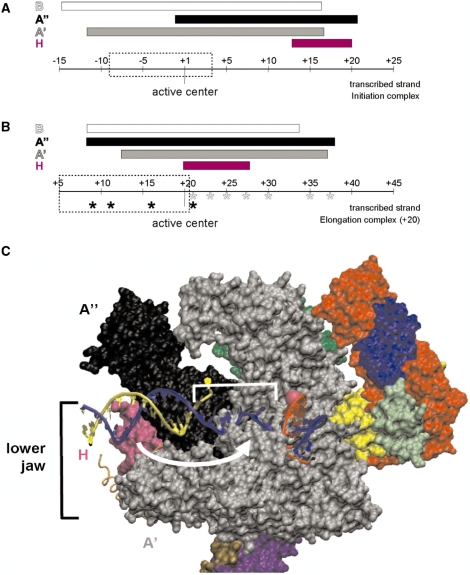
The lower jaws of archaeal RNA polymerase and eukaryotic RNA polymerase II include orthologous subunits H and Rpb5, respectively. The tertiary structure of H is very similar to the structure of the C-terminal domain of Rpb5, and both subunits are proximal to downstream DNA in pre-initiation complexes. Analyses of reconstituted euryarchaeal polymerase lacking subunit H revealed that H is important for open complex formation and initial transcription. Eukaryotic Rpb5 rescues activity of the DeltaH enzyme indicating a strong conservation of function for this subunit from archaea to eukaryotes. Photochemical cross-linking in elongation complexes revealed a striking structural rearrangement of RNA polymerase, bringing subunit H near the transcribed DNA strand one helical turn downstream of the active center, in contrast to the positioning observed in preinitiation complexes. The rearrangement of subunits H and A'' suggest a major conformational change in the archaeal RNAP lower jaw upon formation of the elongation complex.
Figures
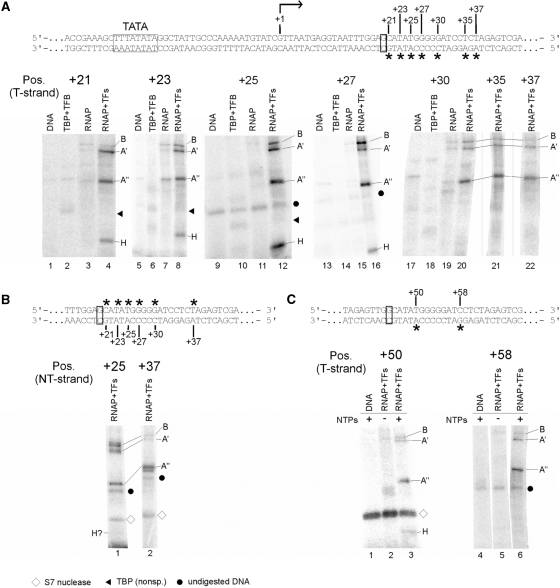
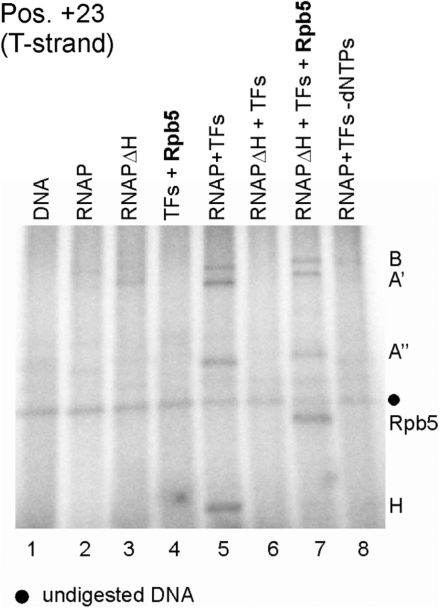
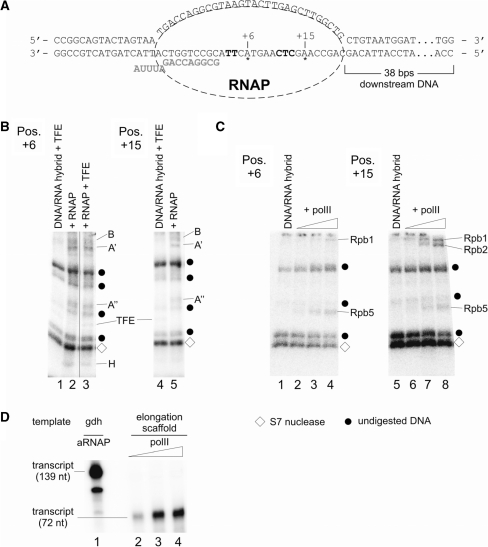
References
-
- Hausner W, Wettach J, Hethke C, Thomm M. Two transcription factors related with the eucaryal transcription factors TATA-binding protein and transcription factor IIB direct promoter recognition by an archaeal RNA polymerase. J. Biol. Chem. 1996;271:30144–30148. - PubMed
-
- Bell SD, Jackson P. The role of transcription factor B in transcription initiation and promoter clearance in the Archaeon Sulfolobus acidocaldarius. J. Biol. Chem. 2000;275:12934–12940. - PubMed
-
- Meinhart AJ, Blobel J, Cramer P. An extended winged helix domain in general transcription factor E/IIEalpha. J. Biol. Chem. 2003;278:48267–48274. - PubMed
