

Multiple pathways ensure retinoid delivery to milk: studies in genetically modified mice
- PMID: 20040693
- PMCID: PMC2853214
- DOI: 10.1152/ajpendo.00491.2009
Multiple pathways ensure retinoid delivery to milk: studies in genetically modified mice
Abstract
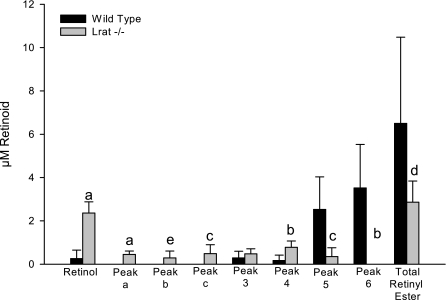
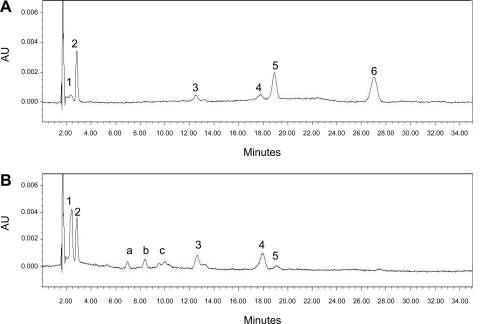
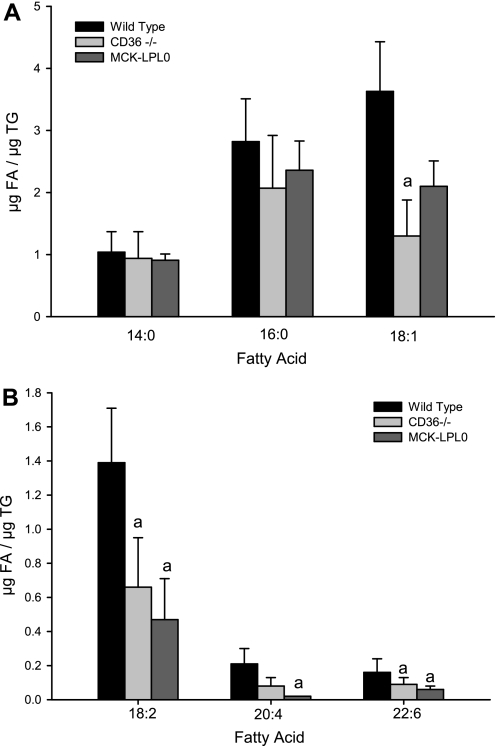
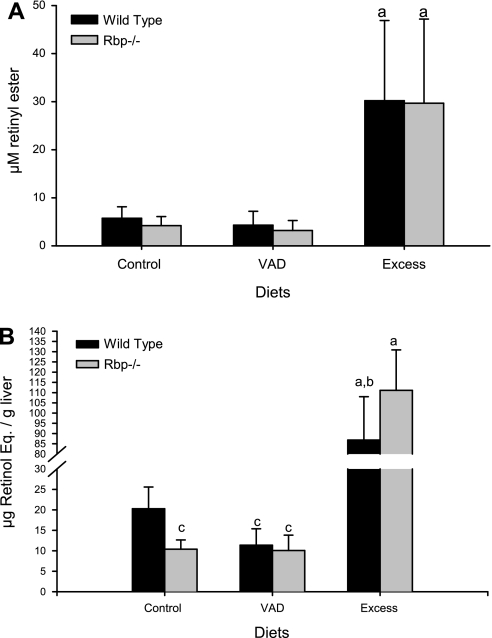
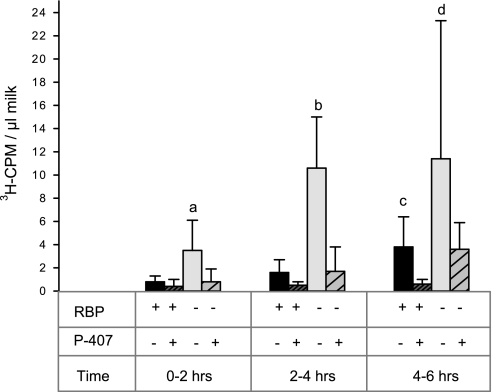
Retinoids are absolutely required for normal growth and development during the postnatal period. We studied the delivery of retinoids to milk, availing of mouse models modified for proteins thought to be essential for this process. Milk retinyl esters were markedly altered in mice lacking the enzyme lecithin:retinol acyltransferase (Lrat(-/-)), indicating that this enzyme is normally responsible for the majority of retinyl esters incorporated into milk and not an acyl-CoA dependent enzyme, as proposed in the literature. Unlike wild-type milk, much of the retinoid in Lrat(-/-) milk is unesterified retinol, not retinyl ester. The composition of the residual retinyl ester present in Lrat(-/-) milk was altered from predominantly retinyl palmitate and stearate to retinyl oleate and medium chain retinyl esters. This was accompanied by increased palmitate and decreased oleate in Lrat(-/-) milk triglycerides. In other studies, we investigated the role of retinol-binding protein in retinoid delivery for milk formation. We found that Rbp(-/-) mice maintain milk retinoid concentrations similar to those in matched wild-type mice. This appears to arise due to greater postprandial delivery of retinoid, a lipoprotein lipase (LPL)-dependent pathway. Importantly, LPL also acts to assure delivery of long-chain fatty acids (LCFA) to milk. The fatty acid transporter CD36 also facilitated LCFA but not retinoid incorporation into milk. Our data show that compensatory pathways for the delivery of retinoids ensure their optimal delivery and that LRAT is the most important enzyme for milk retinyl ester formation.
Figures
References
-
- Balmer JE, Blomhoff R. Gene expression regulation by retinoic acid. J Lipid Res 43: 1773–1808, 2002 - PubMed
-
- Berger GM, Spark A, Baillie PM, Huskisson J, Stockwell G, van der Merwe E. Absence of serum-stimulated lipase activity and altered lipid content in milk from a patient with type I hyperlipoproteinaemia. Pediatr Res 17: 835–839, 1983 - PubMed
-
- Blaner WS, Das SR, Gouras P, Flood MT. Hydrolysis of 11-cis- and all-trans-retinyl palmitate by homogenates of human retinal epithelial cells. J Biol Chem 262: 53–58, 1987. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases