

Bacterial effector binding to ribosomal protein s3 subverts NF-kappaB function
- PMID: 20041225
- PMCID: PMC2791202
- DOI: 10.1371/journal.ppat.1000708
Bacterial effector binding to ribosomal protein s3 subverts NF-kappaB function
Abstract
Enteric bacterial pathogens cause food borne disease, which constitutes an enormous economic and health burden. Enterohemorrhagic Escherichia coli (EHEC) causes a severe bloody diarrhea following transmission to humans through various means, including contaminated beef and vegetable products, water, or through contact with animals. EHEC also causes a potentially fatal kidney disease (hemolytic uremic syndrome) for which there is no effective treatment or prophylaxis. EHEC and other enteric pathogens (e.g., enteropathogenic E. coli (EPEC), Salmonella, Shigella, Yersinia) utilize a type III secretion system (T3SS) to inject virulence proteins (effectors) into host cells. While it is known that T3SS effectors subvert host cell function to promote diarrheal disease and bacterial transmission, in many cases, the mechanisms by which these effectors bind to host proteins and disrupt the normal function of intestinal epithelial cells have not been completely characterized. In this study, we present evidence that the E. coli O157:H7 nleH1 and nleH2 genes encode T3SS effectors that bind to the human ribosomal protein S3 (RPS3), a subunit of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kappaB) transcriptional complexes. NleH1 and NleH2 co-localized with RPS3 in the cytoplasm, but not in cell nuclei. The N-terminal region of both NleH1 and NleH2 was required for binding to the N-terminus of RPS3. NleH1 and NleH2 are autophosphorylated Ser/Thr protein kinases, but their binding to RPS3 is independent of kinase activity. NleH1, but not NleH2, reduced the nuclear abundance of RPS3 without altering the p50 or p65 NF-kappaB subunits or affecting the phosphorylation state or abundance of the inhibitory NF-kappaB chaperone IkappaBalpha NleH1 repressed the transcription of a RPS3/NF-kappaB-dependent reporter plasmid, but did not inhibit the transcription of RPS3-independent reporters. In contrast, NleH2 stimulated RPS3-dependent transcription, as well as an AP-1-dependent reporter. We identified a region of NleH1 (N40-K45) that is at least partially responsible for the inhibitory activity of NleH1 toward RPS3. Deleting nleH1 from E. coli O157:H7 produced a hypervirulent phenotype in a gnotobiotic piglet model of Shiga toxin-producing E. coli infection. We suggest that NleH may disrupt host innate immune responses by binding to a cofactor of host transcriptional complexes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
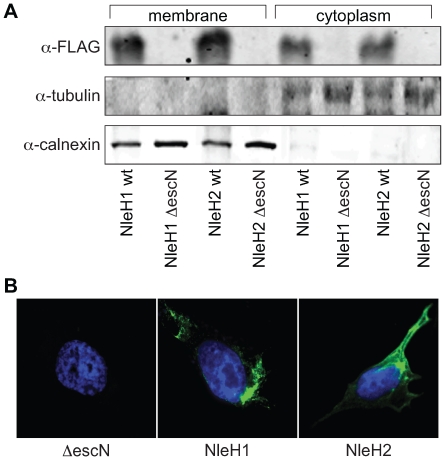
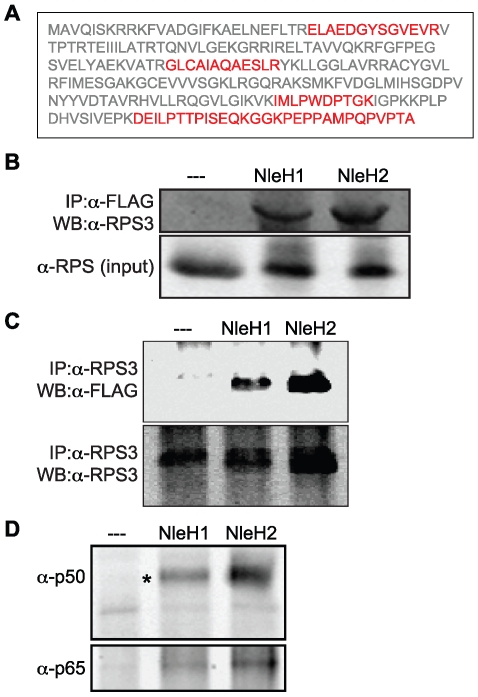
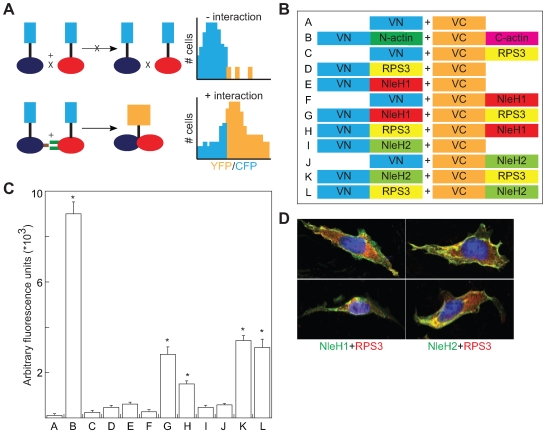
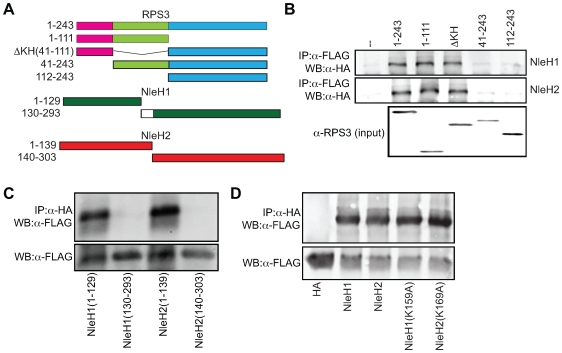
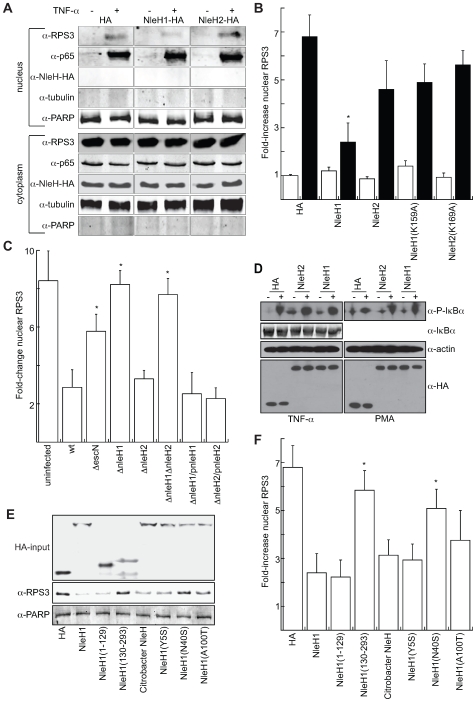
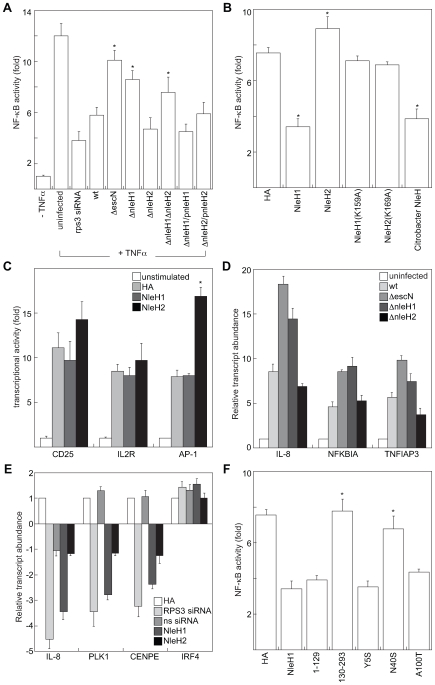
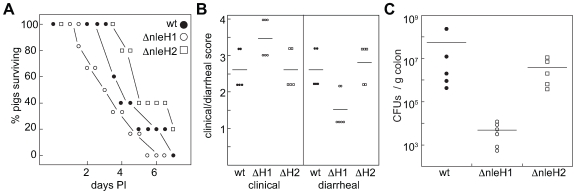
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
