

Echinoderms: potential model systems for studies on muscle regeneration
- PMID: 20041824
- PMCID: PMC2933377
- DOI: 10.2174/138161210790883426
Echinoderms: potential model systems for studies on muscle regeneration
Abstract
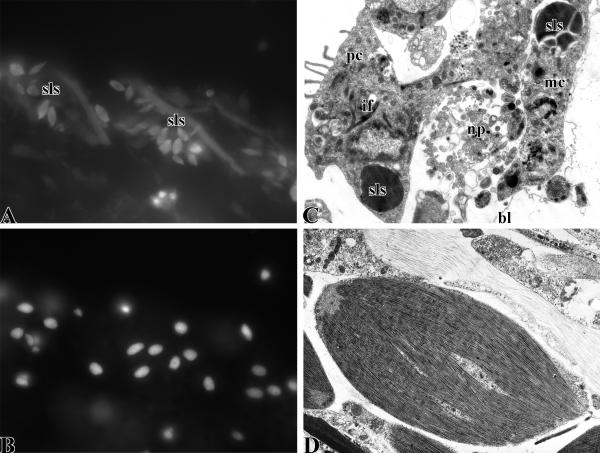
Organisms of the phylum Echinodermata show some of the most impressive regenerative feats within the animal kingdom. Following injury or self-induced autotomy, species in this phylum can regenerate most tissues and organs, being the regeneration of the muscular systems one of the best studied. Even though echinoderms are closely related to chordates, they are little known in the biomedical field, and therefore their uses to study pharmacological effects on muscle formation and/or regeneration have been extremely limited. In order to rectify this lack of knowledge, we describe here the echinoderm muscular systems, particularly the somatic and visceral muscle components. In addition, we provide details of the processes that are known to take place during muscle regeneration, namely dedifferentiation, myogenesis and new muscle formation. Finally, we provide the available information on molecular and pharmacological studies that involve echinoderm muscle regeneration. We expect that by making this information accessible, researchers consider the use of echinoderms as model systems for pharmacological studies in muscle development and regeneration.
Figures








References
-
- Sánchez Alvarado A, Tsonis PA. Bridging the regeneration gap: genetic insight from diverse animal models. Nature Rev Genet. 2006;7:873–83. - PubMed
-
- Brockes JP, Kumar A. Comparative aspects of animal regeneration. Annu Rev Cell Dev Biol. 2008;24:525–49. - PubMed
-
- Blair JE, SE, Hedges S. Molecular phylogeny and divergence times of deuterostome animals. Mol Biol Evol. 2005;22:22–75-84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
