

Effect of mutations in the human immunodeficiency virus type 1 protease on cleavage of the gp41 cytoplasmic tail
- PMID: 20042499
- PMCID: PMC2826043
- DOI: 10.1128/JVI.02002-09
Effect of mutations in the human immunodeficiency virus type 1 protease on cleavage of the gp41 cytoplasmic tail
Abstract
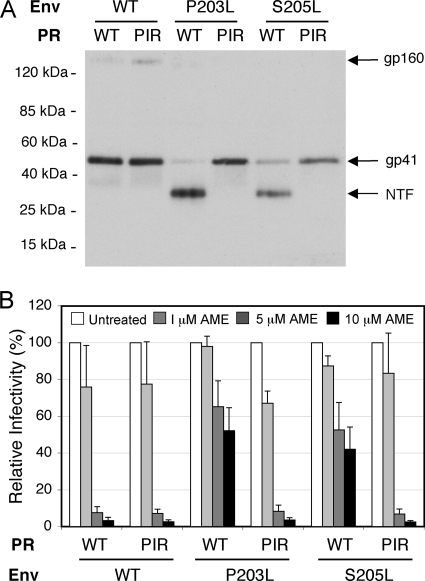
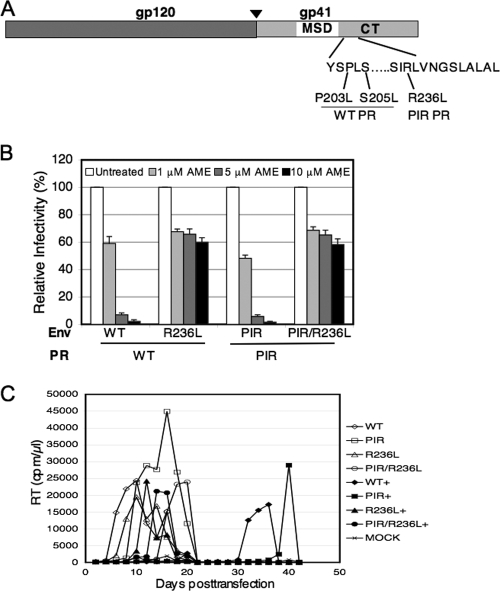
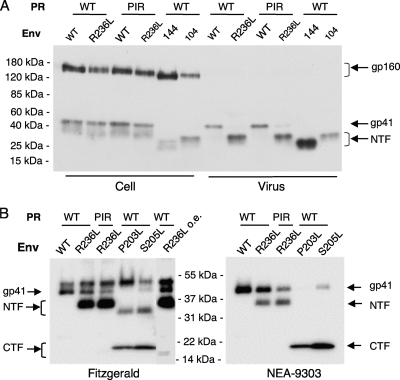
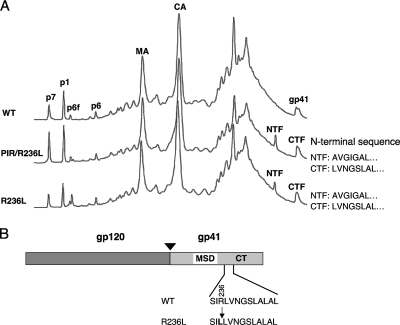
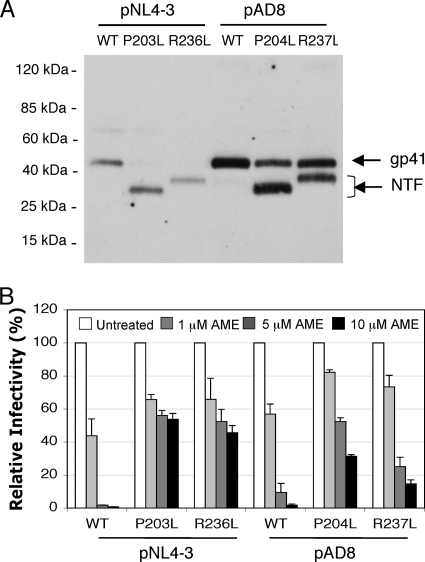
We previously reported that human immunodeficiency virus type 1 (HIV-1) develops resistance to the cholesterol-binding compound amphotericin B methyl ester (AME) by acquiring mutations (P203L and S205L) in the cytoplasmic tail of the transmembrane envelope glycoprotein gp41 that create cleavage sites for the viral protease (PR). In the present study, we observed that a PR inhibitor-resistant (PIR) HIV-1 mutant is unable to efficiently cleave the gp41 cytoplasmic tail in P203L and S205L virions, resulting in loss of AME resistance. To define the pathway to AME resistance in the context of the PIR PR, we selected for resistance with an HIV-1 isolate expressing the mutant enzyme. We identified a new gp41 mutation, R236L, that results in cleavage of the gp41 tail by the PIR PR. These results highlight the central role of gp41 cleavage as the primary mechanism of AME resistance.
Figures
Similar articles
-
HIV-1 escape from the entry-inhibiting effects of a cholesterol-binding compound via cleavage of gp41 by the viral protease.Proc Natl Acad Sci U S A. 2007 May 15;104(20):8467-71. doi: 10.1073/pnas.0701443104. Epub 2007 May 4. Proc Natl Acad Sci U S A. 2007. PMID: 17483482 Free PMC article.
-
Inhibition of HIV-1 replication by amphotericin B methyl ester: selection for resistant variants.J Biol Chem. 2006 Sep 29;281(39):28699-711. doi: 10.1074/jbc.M603609200. Epub 2006 Aug 1. J Biol Chem. 2006. PMID: 16882663
-
HIV-1 protease, Gag and gp41 baseline substitutions associated with virological response to a PI-based regimen.J Antimicrob Chemother. 2019 Jun 1;74(6):1679-1692. doi: 10.1093/jac/dkz043. J Antimicrob Chemother. 2019. PMID: 30768160
-
Resilience to resistance of HIV-1 protease inhibitors: profile of darunavir.AIDS Rev. 2008 Jul-Sep;10(3):131-42. AIDS Rev. 2008. PMID: 18820715 Free PMC article. Review.
-
Reviewing HIV-1 Gag Mutations in Protease Inhibitors Resistance: Insights for Possible Novel Gag Inhibitor Designs.Molecules. 2019 Sep 6;24(18):3243. doi: 10.3390/molecules24183243. Molecules. 2019. PMID: 31489889 Free PMC article. Review.
Cited by
-
Ceramide synthase 2 deletion decreases the infectivity of HIV-1.J Biol Chem. 2021 Jan-Jun;296:100340. doi: 10.1016/j.jbc.2021.100340. Epub 2021 Jan 28. J Biol Chem. 2021. PMID: 33515546 Free PMC article.
-
The frantic play of the concealed HIV envelope cytoplasmic tail.Retrovirology. 2013 May 24;10:54. doi: 10.1186/1742-4690-10-54. Retrovirology. 2013. PMID: 23705972 Free PMC article. Review.
-
Analysis of HIV-1 envelope cytoplasmic tail effects on viral replication.Virology. 2023 Feb;579:54-66. doi: 10.1016/j.virol.2022.12.017. Epub 2023 Jan 2. Virology. 2023. PMID: 36603533 Free PMC article.
-
HIV Genome-Wide Protein Associations: a Review of 30 Years of Research.Microbiol Mol Biol Rev. 2016 Jun 29;80(3):679-731. doi: 10.1128/MMBR.00065-15. Print 2016 Sep. Microbiol Mol Biol Rev. 2016. PMID: 27357278 Free PMC article.
-
Biochemistry and biophysics of HIV-1 gp41 - membrane interactions and implications for HIV-1 envelope protein mediated viral-cell fusion and fusion inhibitor design.Curr Top Med Chem. 2011 Dec;11(24):2959-84. doi: 10.2174/156802611798808497. Curr Top Med Chem. 2011. PMID: 22044229 Free PMC article. Review.
References
-
- Brown, D. A., and E. London. 2000. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 275:17221-17224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
