

Mechanisms of apoptosis in Crustacea: What conditions induce versus suppress cell death?
- PMID: 20043212
- PMCID: PMC4104421
- DOI: 10.1007/s10495-009-0443-6
Mechanisms of apoptosis in Crustacea: What conditions induce versus suppress cell death?
Abstract
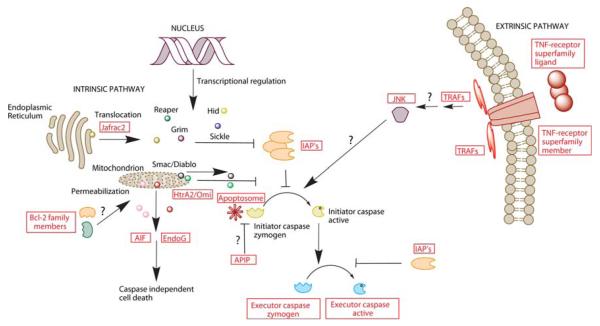
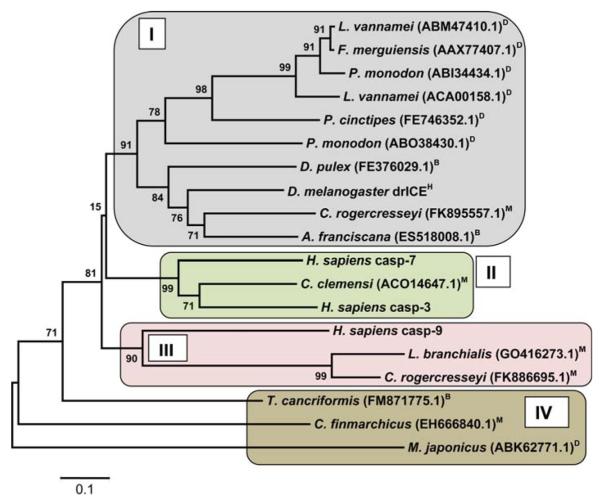
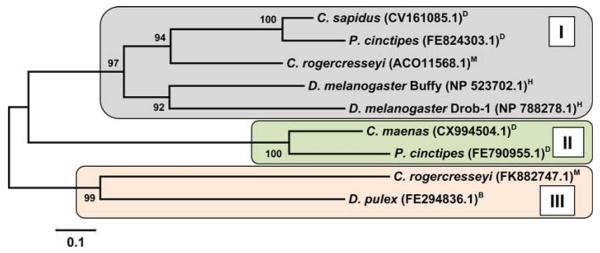
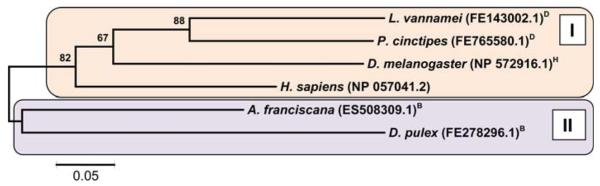
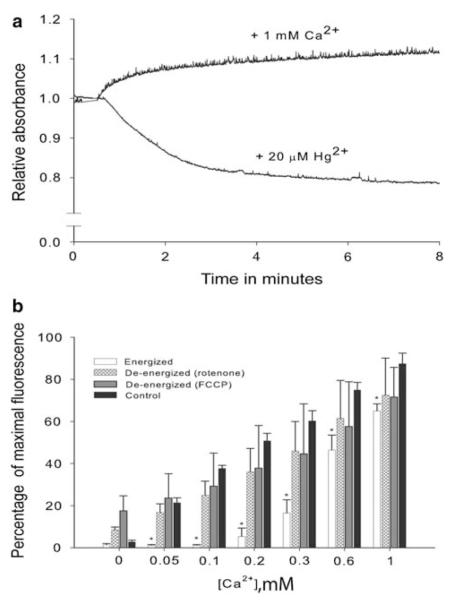
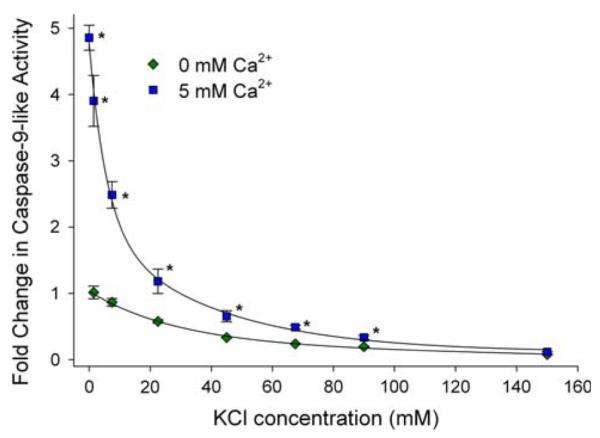
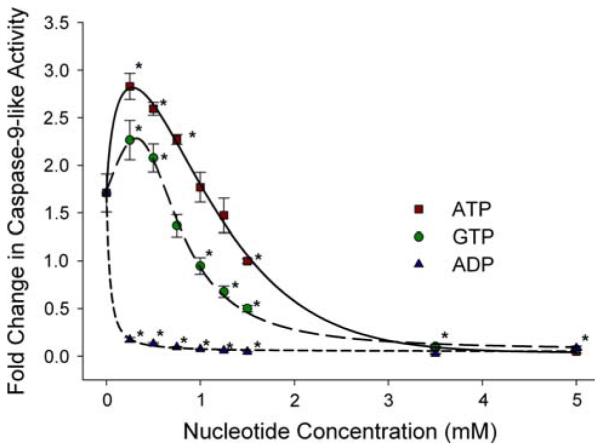
Arthropoda is the largest of all animal phyla and includes about 90% of extant species. Our knowledge about regulation of apoptosis in this phylum is largely based on findings for the fruit fly Drosophila melanogaster. Recent work with crustaceans shows that apoptotic proteins, and presumably mechanisms of cell death regulation, are more diverse in arthropods than appreciated based solely on the excellent work with fruit flies. Crustacean homologs exist for many major proteins in the apoptotic networks of mammals and D. melanogaster, but integration of these proteins into the physiology and pathophysiology of crustaceans is far from complete. Whether apoptosis in crustaceans is mainly transcriptionally regulated as in D. melanogaster (e.g., RHG 'killer' proteins), or rather is controlled by pro- and anti-apoptotic Bcl-2 family proteins as in vertebrates needs to be clarified. Some phenomena like the calcium-induced opening of the mitochondrial permeability transition pore (MPTP) are apparently lacking in crustaceans and may represent a vertebrate invention. We speculate that differences in regulation of the intrinsic pathway of crustacean apoptosis might represent a prerequisite for some species to survive harsh environmental insults. Pro-apoptotic stimuli described for crustaceans include UV radiation, environmental toxins, and a diatom-produced chemical that promotes apoptosis in offspring of a copepod. Mechanisms that serve to depress apoptosis include the inhibition of caspase activity by high potassium in energetically healthy cells, alterations in nucleotide abundance during energy-limited states like diapause and anoxia, resistance to opening of the calcium-induced MPTP, and viral accommodation during persistent viral infection. Characterization of the players, pathways, and their significance in the core machinery of crustacean apoptosis is revealing new insights for the field of cell death.
Figures

References
-
- Danial NN, Korsmeyer SJ. Cell death: critical control points. Cell. 2004;116:205–219. - PubMed
-
- Hassanin A. Phylogeny of Arthropoda inferred from mitochondrial sequences: strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution. Mol Phylogenet Evol. 2006;38:100–116. - PubMed
-
- Bottger A, Alexandrova O. Programmed cell death in Hydra. Semin Cancer Biol. 2007;17:134–146. - PubMed
-
- Robertson AJ, Croce J, Carbonneau S, et al. The genomic underpinnings of apoptosis in Strongylocentrotus purpuratus. Dev Biol. 2006;300:321–334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
