

A substructure combination strategy to create potent and selective transthyretin kinetic stabilizers that prevent amyloidogenesis and cytotoxicity
- PMID: 20043671
- PMCID: PMC2818972
- DOI: 10.1021/ja908562q
A substructure combination strategy to create potent and selective transthyretin kinetic stabilizers that prevent amyloidogenesis and cytotoxicity
Abstract
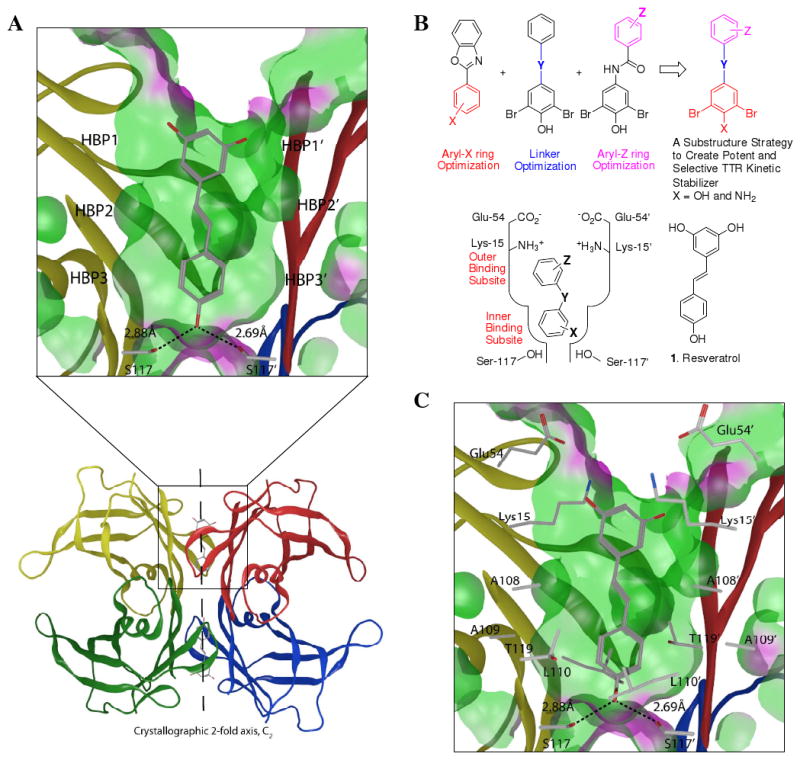
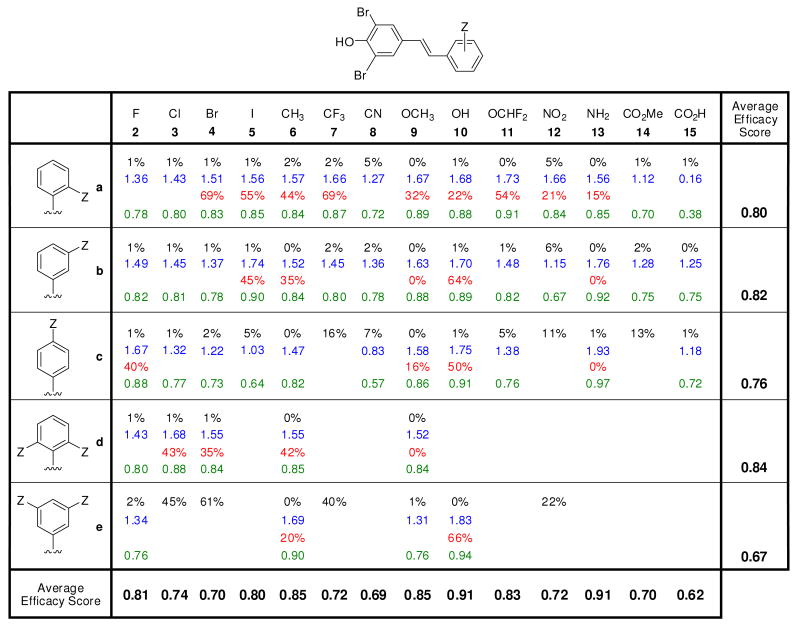
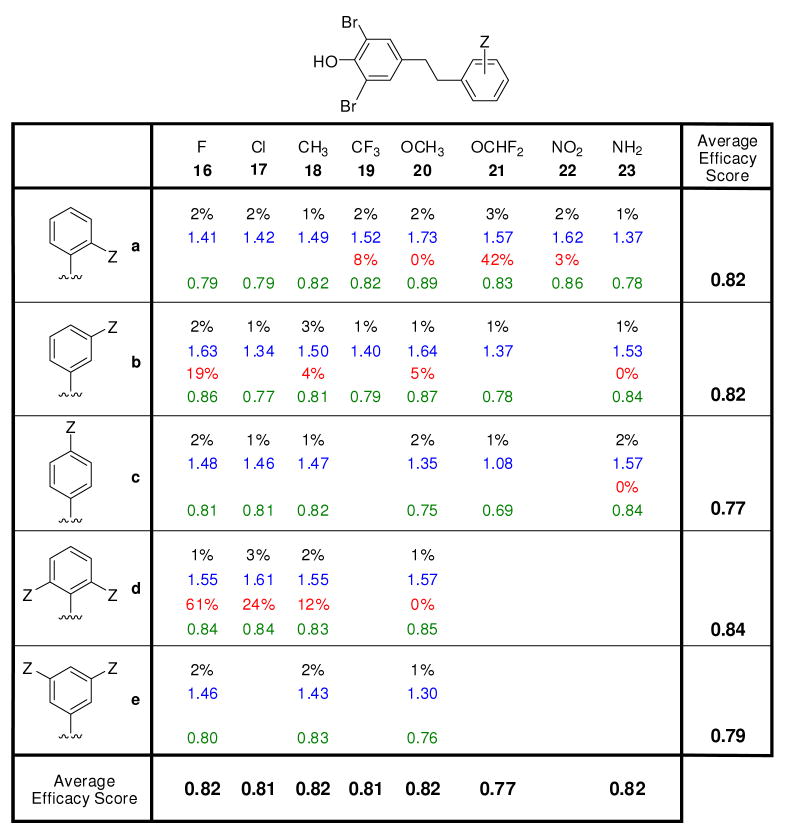
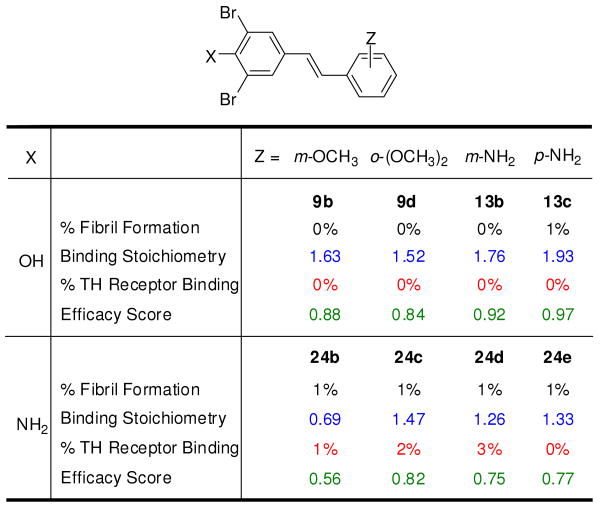
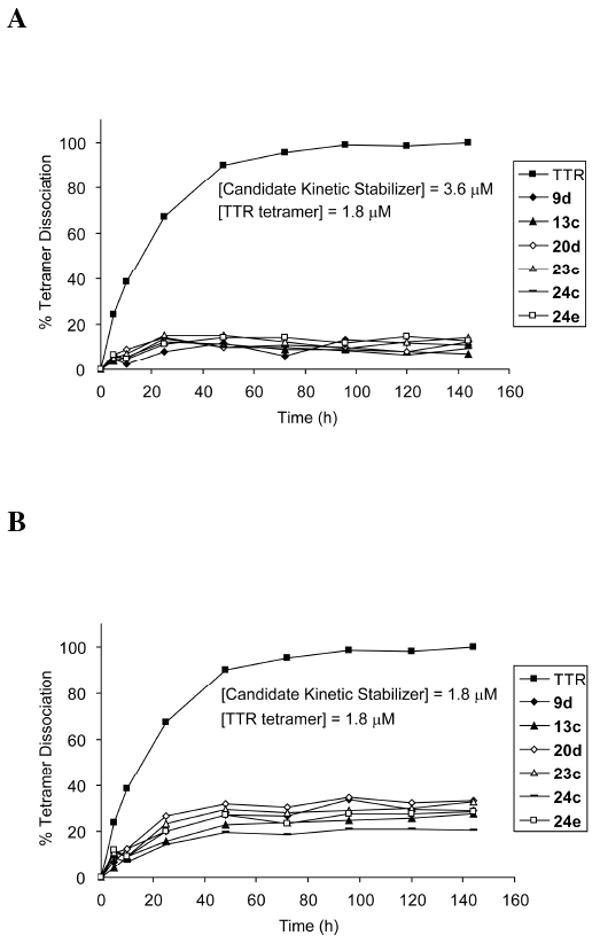
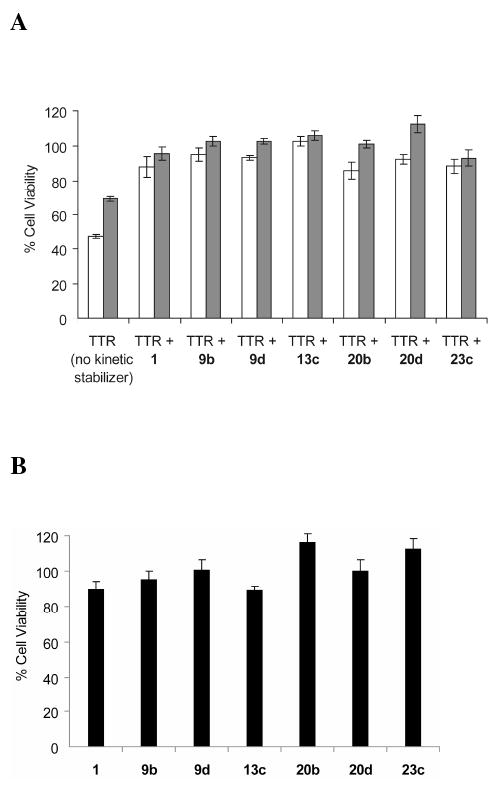
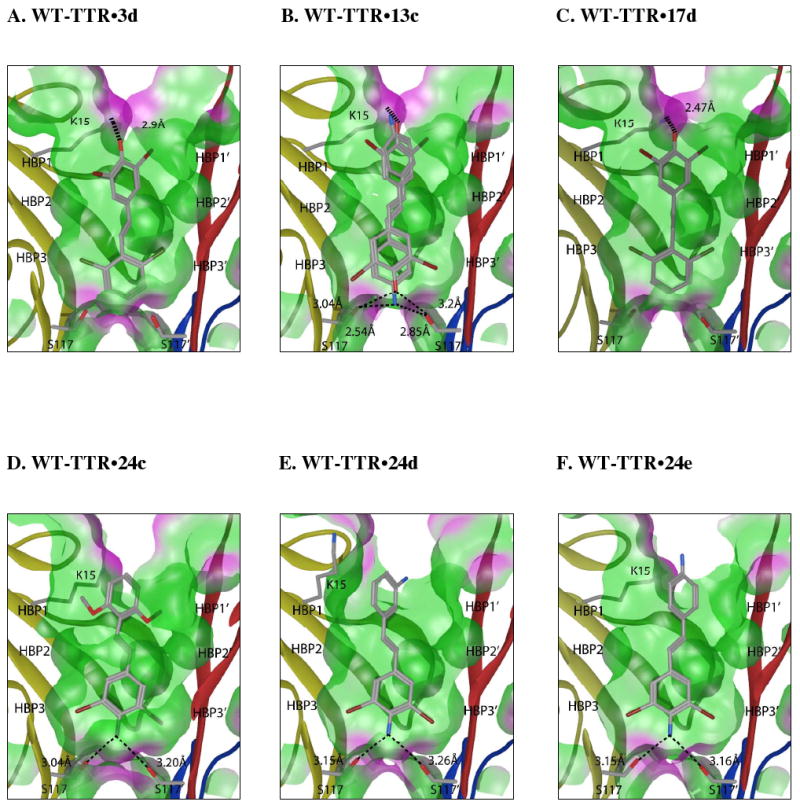
Transthyretin aggregation-associated proteotoxicity appears to cause several human amyloid diseases. Rate-limiting tetramer dissociation and monomer misfolding of transthyretin (TTR) occur before its aggregation into cross-beta-sheet amyloid fibrils. Small molecule binding to and preferential stabilization of the tetrameric state of TTR over the dissociative transition state raises the kinetic barrier for dissociation, imposing kinetic stabilization on TTR and preventing aggregation. This is an effective strategy to halt neurodegeneration associated with polyneuropathy, according to recent placebo-controlled clinical trial results. In three recent papers, we systematically ranked possibilities for the three substructures composing a typical TTR kinetic stabilizer, using fibril inhibition potency and plasma TTR binding selectivity data. Herein, we have successfully employed a substructure combination strategy to use these data to develop potent and selective TTR kinetic stabilizers that rescue cells from the cytotoxic effects of TTR amyloidogenesis. Of the 92 stilbene and dihydrostilbene analogues synthesized, nearly all potently inhibit TTR fibril formation. Seventeen of these exhibit a binding stoichiometry of >1.5 of a maximum of 2 to plasma TTR, while displaying minimal binding to the thyroid hormone receptor (<20%). Six analogues were definitively categorized as kinetic stabilizers by evaluating dissociation time-courses. High-resolution TTR.(kinetic stabilizer)(2) crystal structures (1.31-1.70 A) confirmed the anticipated binding orientation of the 3,5-dibromo-4-hydroxyphenyl substructure and revealed a strong preference of the isosteric 3,5-dibromo-4-aminophenyl substructure to bind to the inner thyroxine binding pocket of TTR.
Figures
References
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Science. 2008;319:916–9. - PubMed
-
- Powers ET, Morimoto RI, Dillin A, Kelly JW, Balch WE. Annu Rev Biochem. 2009;78:959–91. - PubMed
-
- Cohen E, Bieschke J, Perciavalle RM, Kelly JW, Dillin A. Science. 2006;313:1604–10. - PubMed
-
- Sekijima Y, Wiseman RL, Matteson J, Hammarstrom P, Miller SR, Sawkar AR, Balch WE, Kelly JW. Cell. 2005;121:73–85. - PubMed
-
- Hammarstrom P, Wiseman RL, Powers ET, Kelly JW. Science. 2003;299:713–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
