

Nitric oxide synthase-2 regulates mitochondrial Hsp60 chaperone function during bacterial peritonitis in mice
- PMID: 20043987
- PMCID: PMC2842938
- DOI: 10.1016/j.freeradbiomed.2009.12.019
Nitric oxide synthase-2 regulates mitochondrial Hsp60 chaperone function during bacterial peritonitis in mice
Abstract
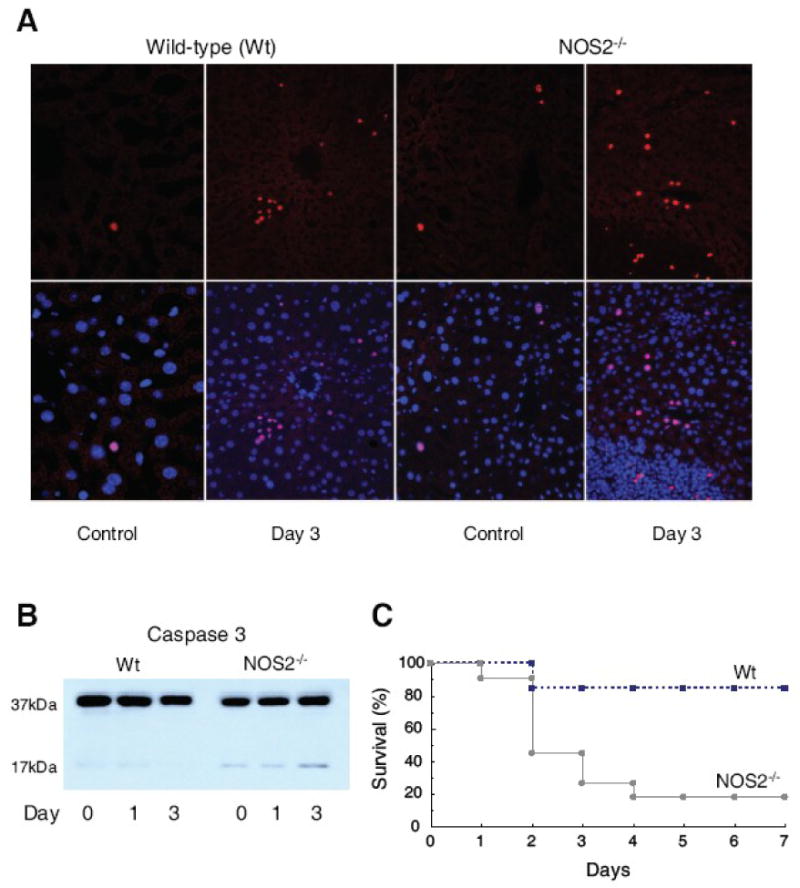
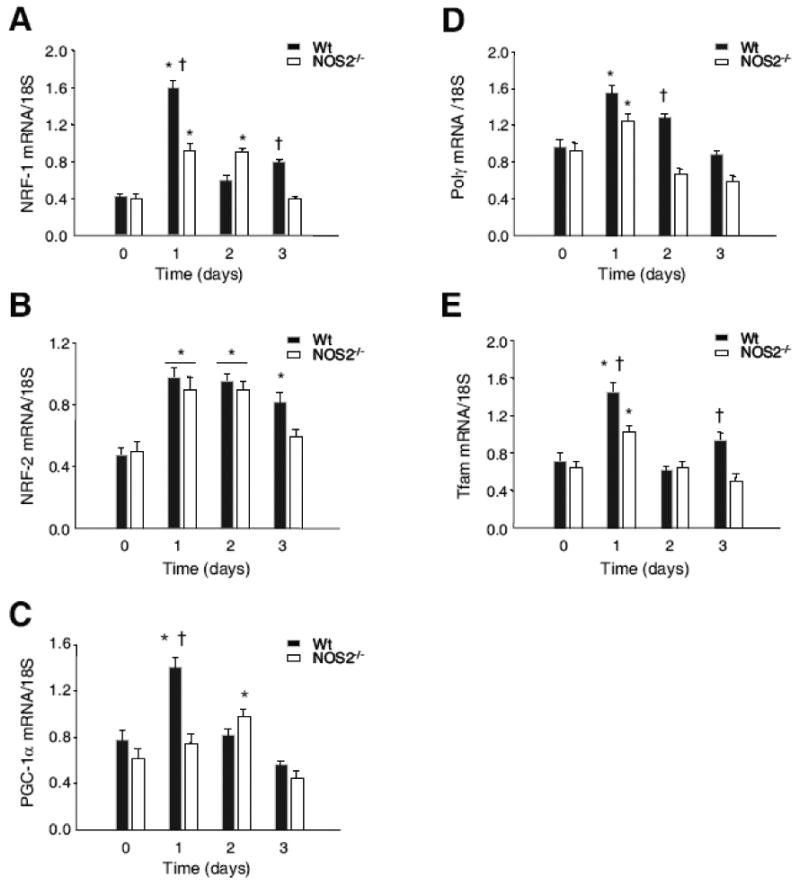
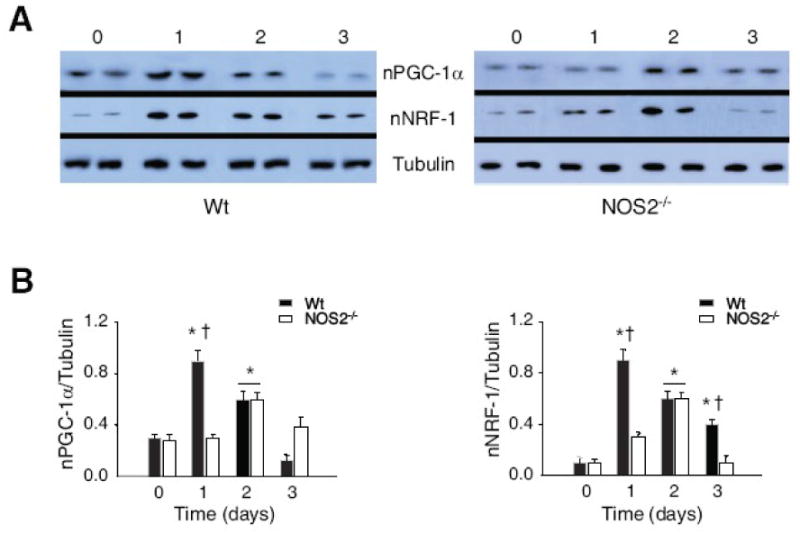
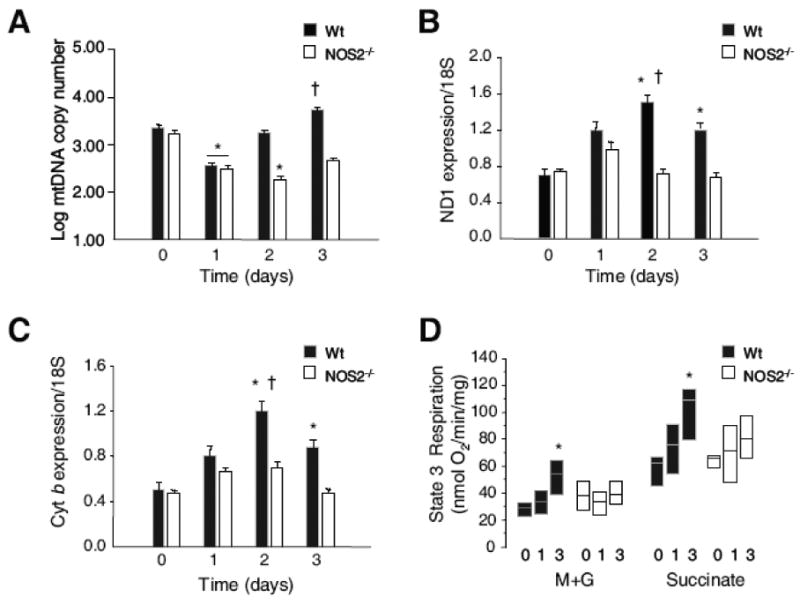
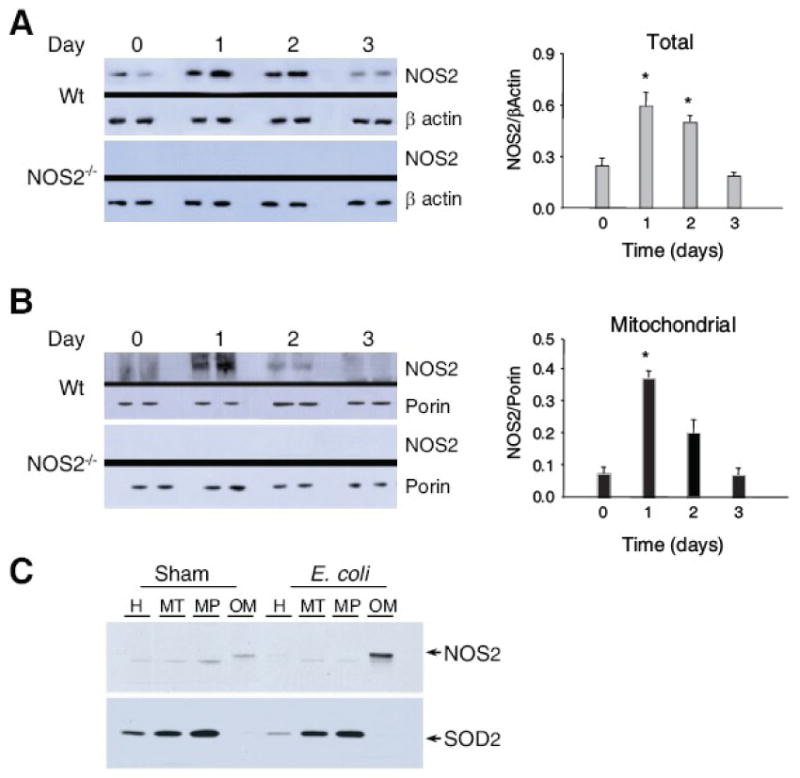
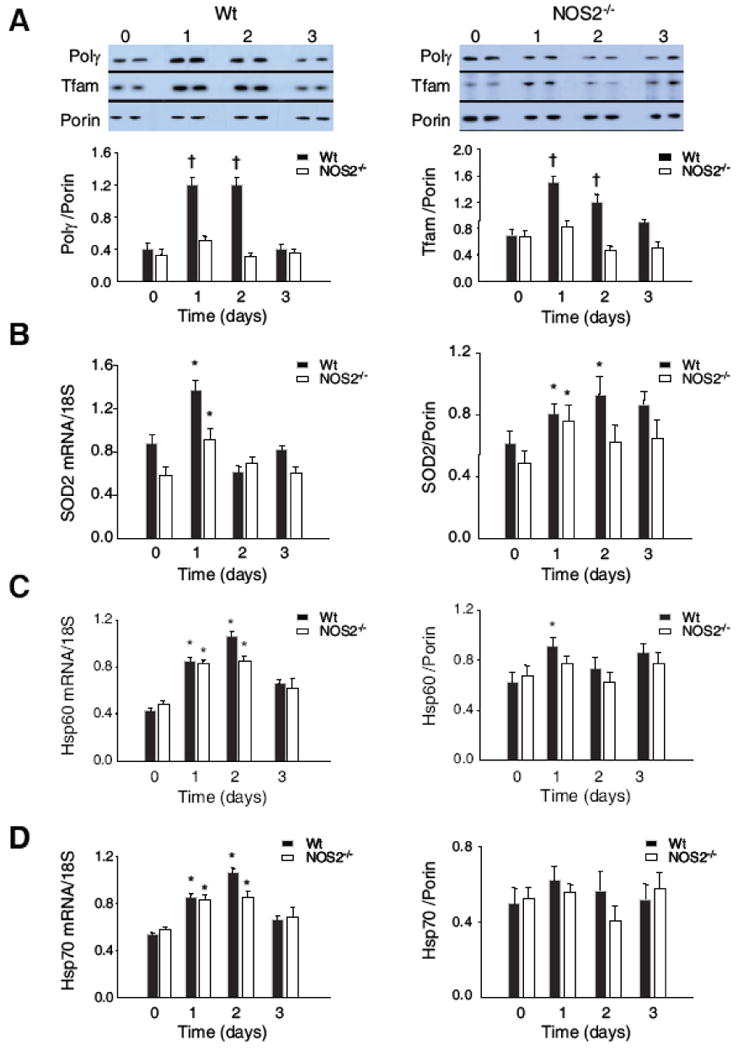
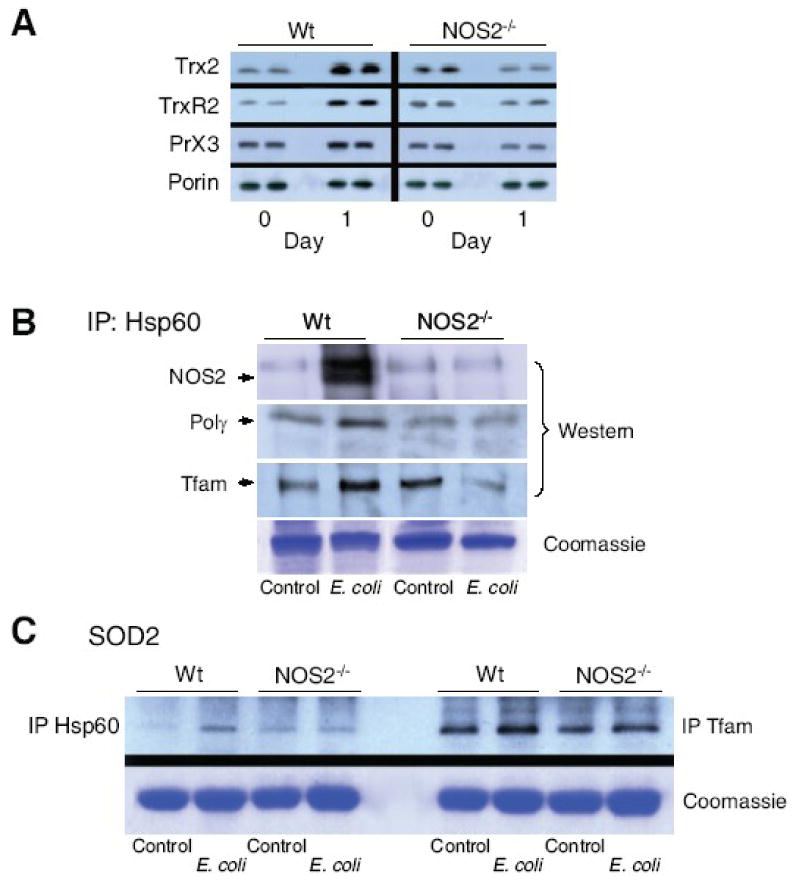
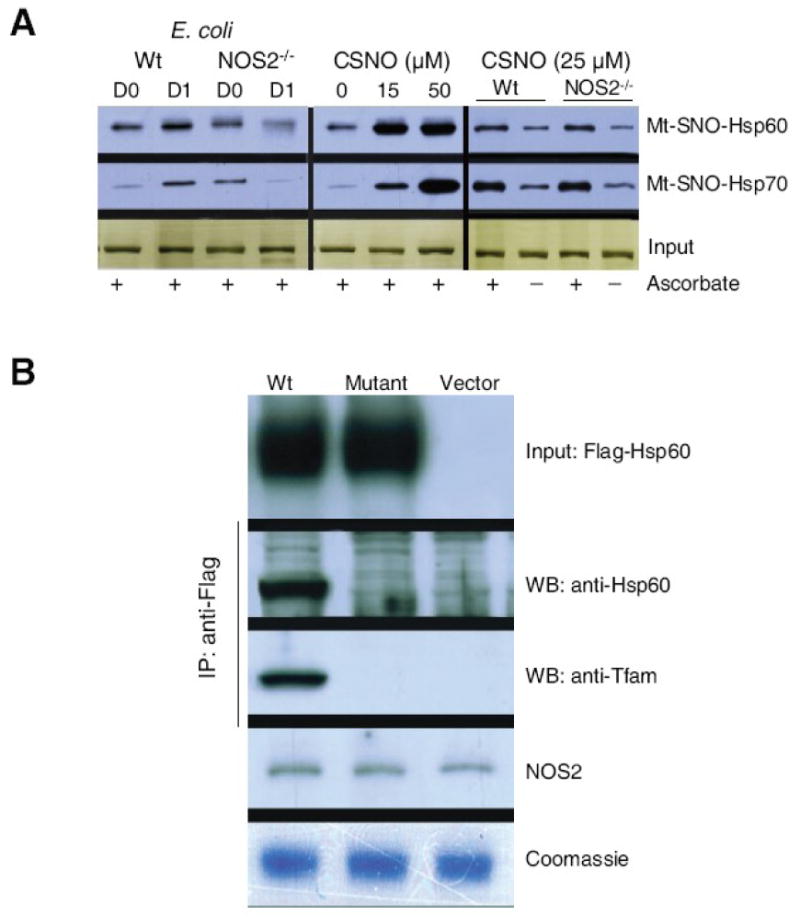
Nitric oxide synthase-2 (NOS2) plays a critical role in reactive nitrogen species generation and cysteine modifications that influence mitochondrial function and signaling during inflammation. Here, we investigated the role of NOS2 in hepatic mitochondrial biogenesis during Escherichia coli peritonitis in mice. NOS2(-/-) mice displayed smaller mitochondrial biogenesis responses than Wt mice during E. coli infection according to differences in mRNA levels for the PGC-1 alpha coactivator, nuclear respiratory factor-1, mitochondrial transcription factor-A (Tfam), and mtDNA polymerase (Pol gamma). NOS2(-/-) mice did not significantly increase mitochondrial Tfam and Pol gamma protein levels during infection in conjunction with impaired mitochondrial DNA (mtDNA) transcription, loss of mtDNA copy number, and lower State 3 respiration rates. NOS2 blockade in mitochondrial-GFP reporter mice disrupted Hsp60 localization to mitochondria after E. coli exposure. Mechanistically, biotin-switch and immunoprecipitation studies demonstrated NOS2 binding to and S-nitros(yl)ation of Hsp60 and Hsp70. Specifically, NOS2 promoted Tfam accumulation in mitochondria by regulation of Hsp60-Tfam binding via S-nitros(yl)ation. In hepatocytes, site-directed mutagenesis identified (237)Cys as a critical residue for Hsp60 S-nitros(yl)ation. Thus, the role of NOS2 in inflammation-induced mitochondrial biogenesis involves both optimal gene expression for nuclear-encoded mtDNA-binding proteins and functional regulation of the Hsp60 chaperone that enables their importation for mtDNA transcription and replication.
(c) 2009 Elsevier Inc. All rights reserved.
Figures
References
-
- Angus DC, Linde-Zwirble WT, Lidicker J, Clermont G, Carcillo J, Pinsky MR. Epidemiology of severe sepsis in the United States: analysis of incidence, outcome, and associated costs of care. Crit Care Med. 2001;29:1303–1310. - PubMed
-
- Levy RM, Prince JM, Billiar TR. Nitric oxide: a clinical primer. Crit Care Med. 2005;33:S492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
