

Cyclooxygenase-2 expression is related to the epithelial-to-mesenchymal transition in human colon cancers
- PMID: 20046424
- PMCID: PMC2796410
- DOI: 10.3349/ymj.2009.50.6.818
Cyclooxygenase-2 expression is related to the epithelial-to-mesenchymal transition in human colon cancers
Abstract
Purpose: Down-regulation of E-cadherin is a hallmark of the epithelial-to-mesenchymal transition (EMT). EMT progression in cancer cells is associated with the loss of certain epithelial markers and the acquisition of a mesenchymal phenotype, as well as migratory activities. Cyclooxygenase-2 (COX-2) expression is associated with tumor invasion and metastasis in colon cancer. This study investigated the relationship between E-cadherin and COX-2 in colon cancer cells and human colon tumors.
Materials and methods: Colon cancer cell lines and immunohistochemistry were used.
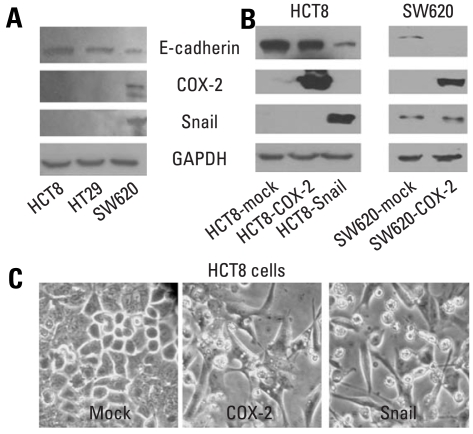
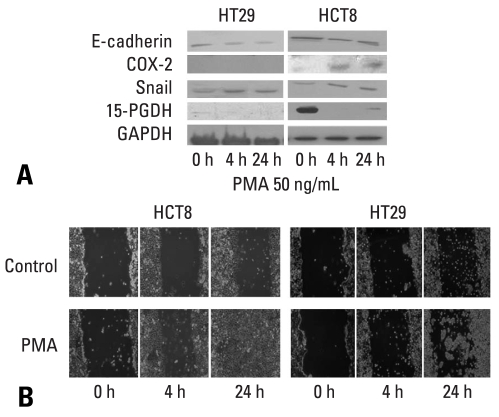
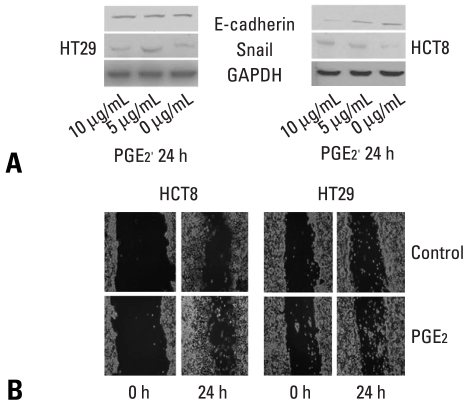
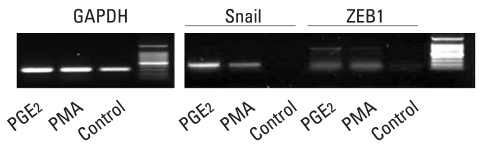
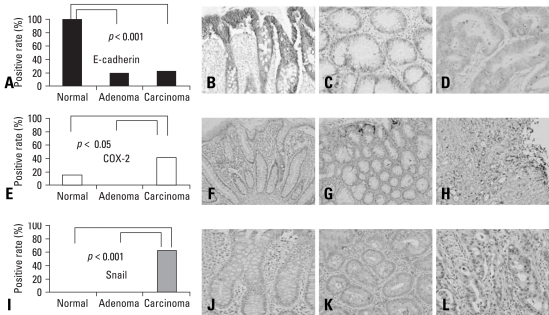
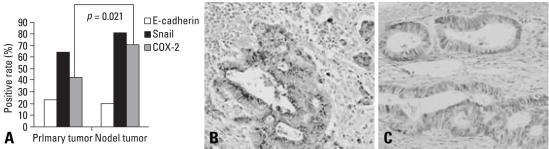
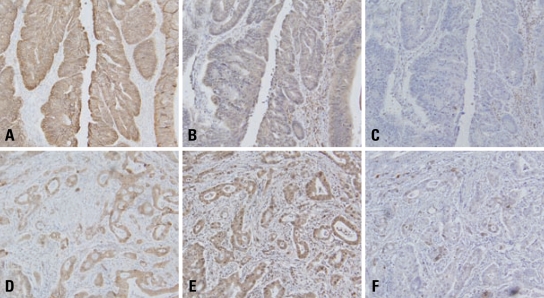
Results: E-cadherin expression was inversely related to the expressions of COX-2 and Snail in colon cancer cells. Ectopic expression of COX-2 or Snail reduced E-cadherin and induced a scattered, flattened phenotype with few intercellular contacts in colon cancer cells. Treatment of cancer cells with phorbol 12-myristate 13-acetate increased the expressions of COX-2 and Snail, decreased 15-hydroxyprostaglandin dehydrogenase expression, and increased the cells' motility. In addition, exposure to prostaglandin E(2) increased Snail expression and cell motility, and decreased E-cadherin expression. Membranous E-cadherin expression was lower in adenomas and cancers than in the adjacent, non-neoplastic epithelium. In contrast, the expressions of Snail and COX-2 were higher in cancers than in normal tissues and adenomas. The expressions of COX-2 and Snail increased in areas with abnormal E-cadherin expression. Moreover, COX-2 expression was related to higher tumor stages and was significantly higher in nodal metastatic lesions than primary cancers.
Conclusion: This study suggests that COX-2 may have a role in tumor metastasis via EMT.
Keywords: COX-2; E-cadherin; Epithelial-to-mesenchymal transition; Snail; colon cancers.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures
References
-
- Brown JR, DuBois RN. COX-2: a molecular target for colorectal cancer prevention. J Clin Oncol. 2005;23:2840–2855. - PubMed
-
- Vleminckx K, Kemler R. Cadherins and tissue formation: integrating adhesion and signaling. Bioessays. 1999;21:211–220. - PubMed
-
- Thiery JP. Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer. 2002;2:442–454. - PubMed
-
- Guarino M, Rubino B, Ballabio G. The role of epithelial-mesenchymal transition in cancer pathology. Pathology. 2007;39:305–318. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
