

Interactions at the 2 and 5 positions of 5-phosphoribosyl pyrophosphate are essential in Salmonella typhimurium quinolinate phosphoribosyltransferase
- PMID: 20047307
- PMCID: PMC2891058
- DOI: 10.1021/bi9018219
Interactions at the 2 and 5 positions of 5-phosphoribosyl pyrophosphate are essential in Salmonella typhimurium quinolinate phosphoribosyltransferase
Abstract
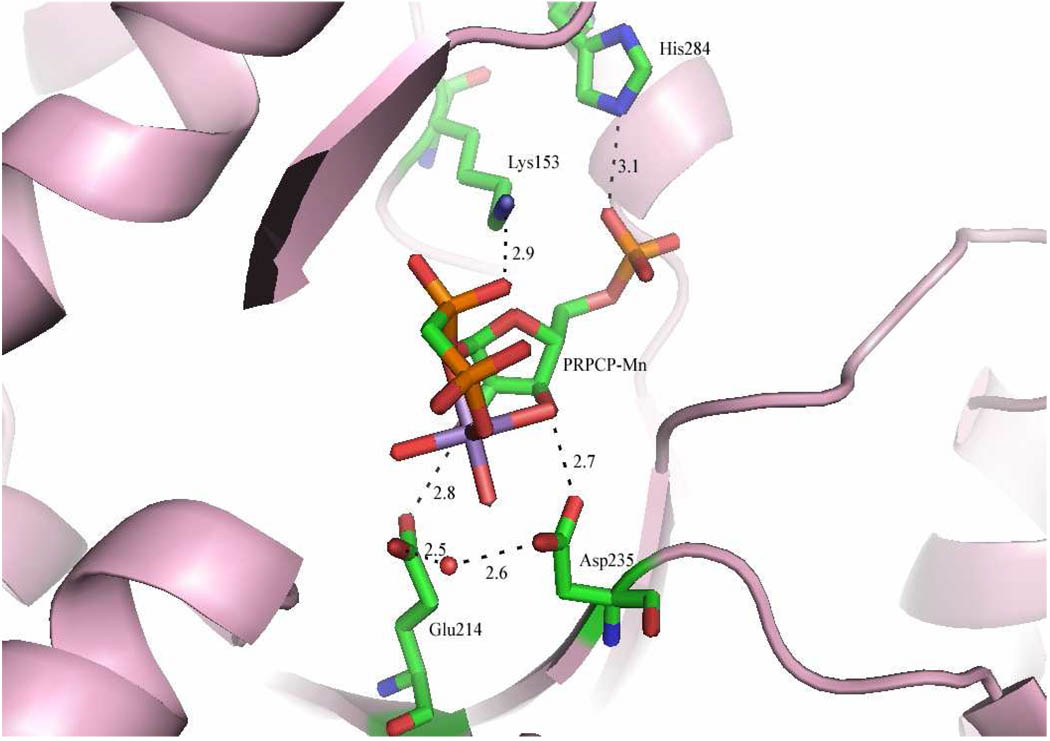
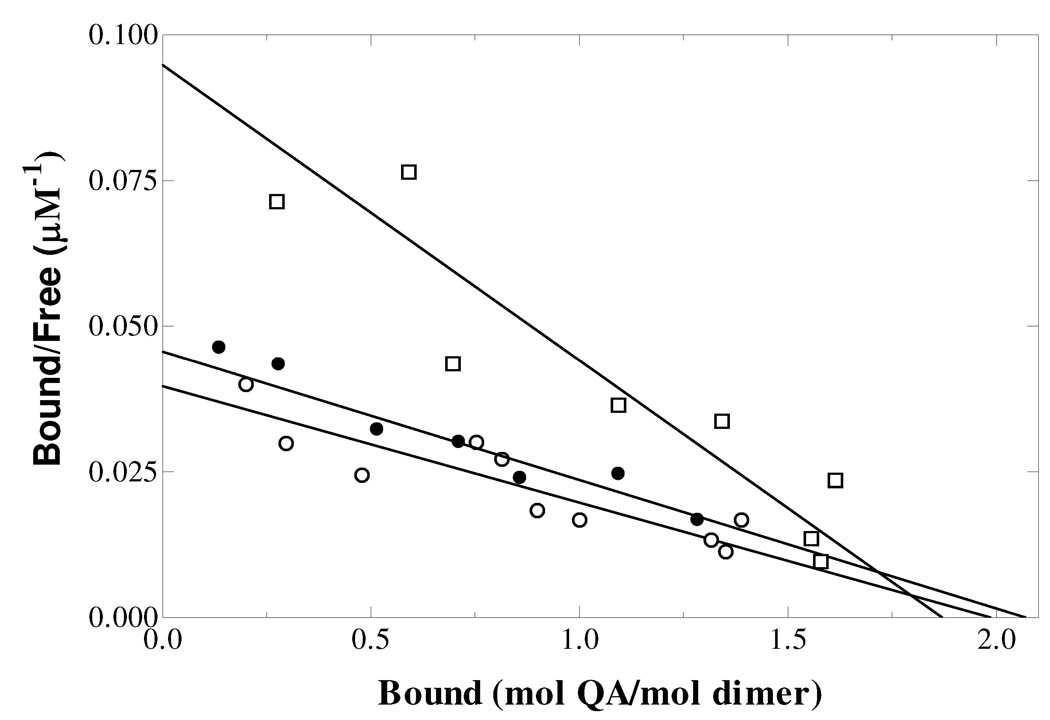
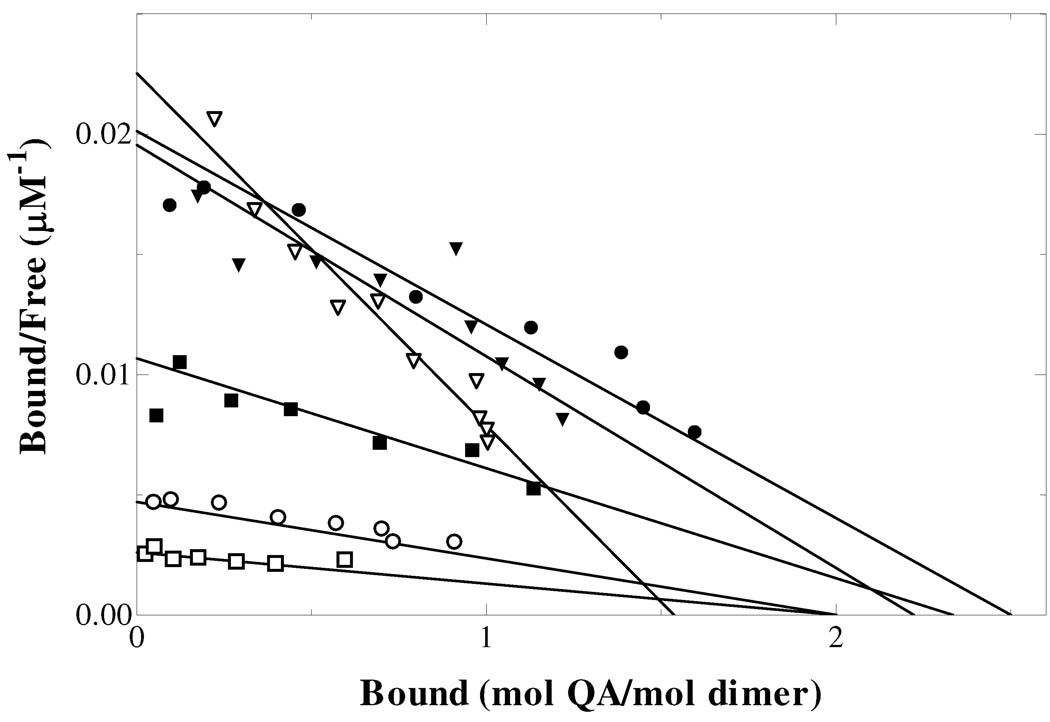
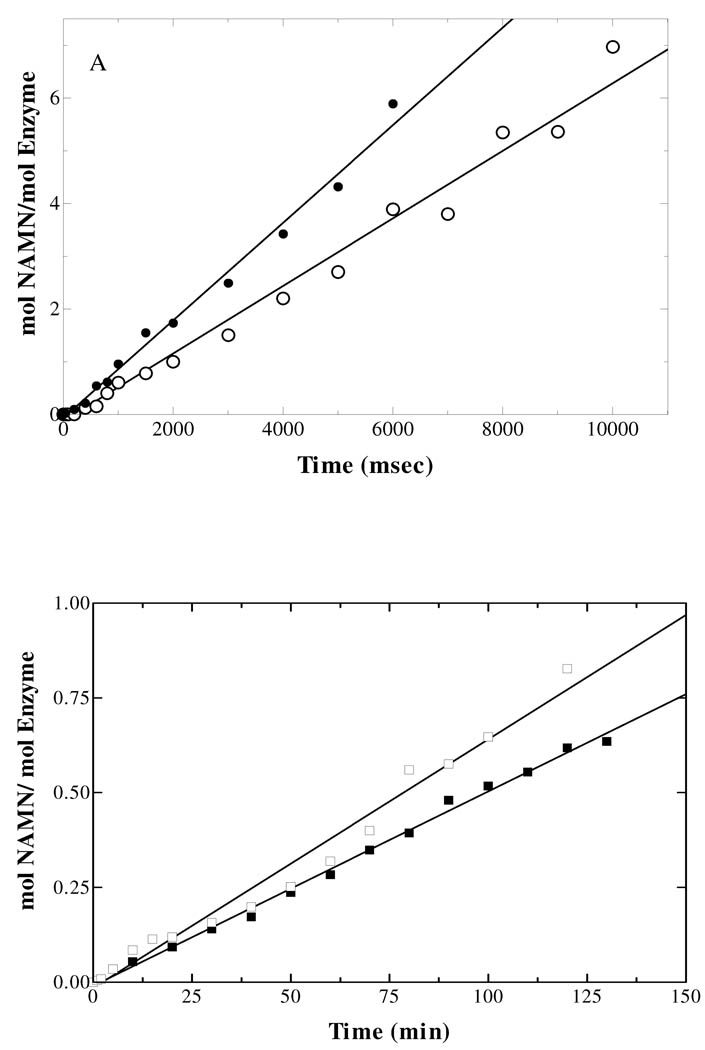
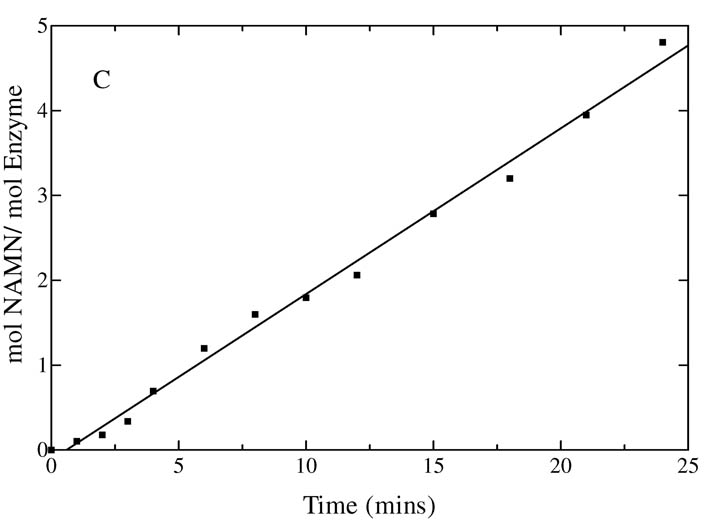
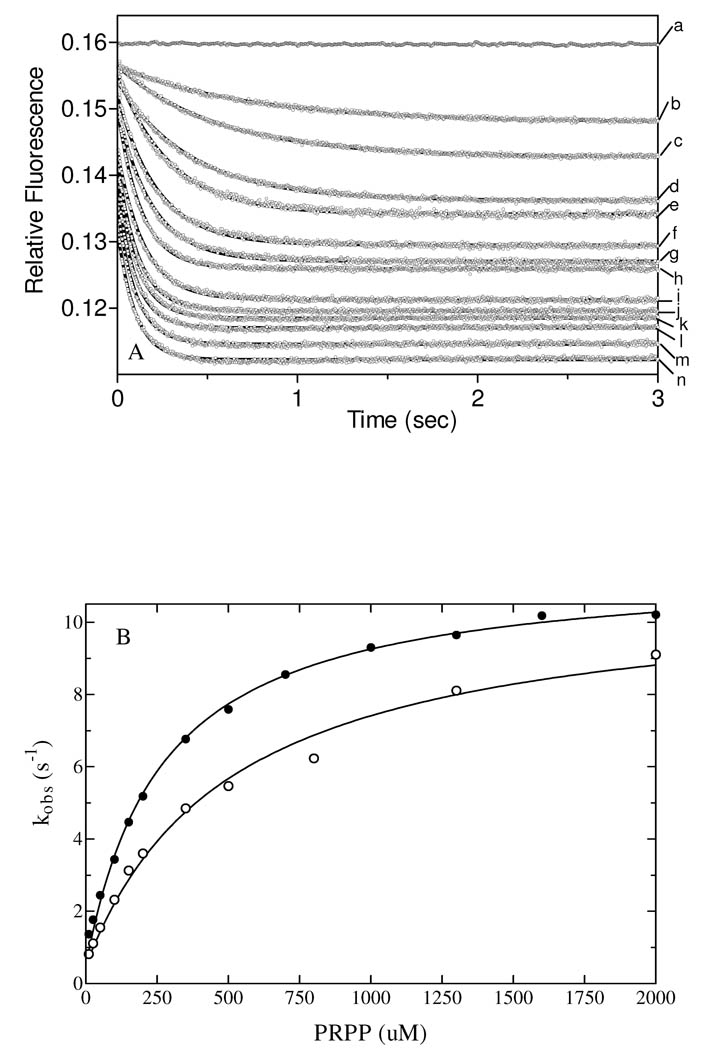
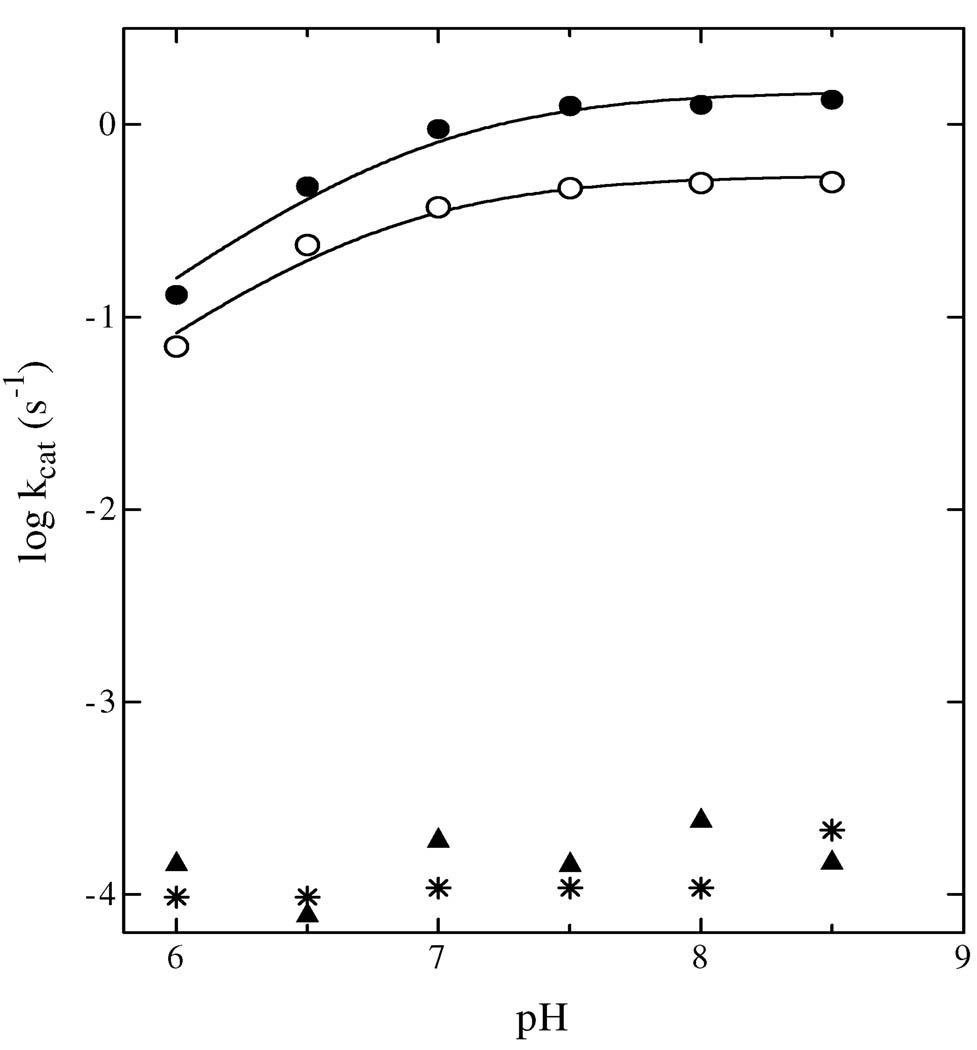
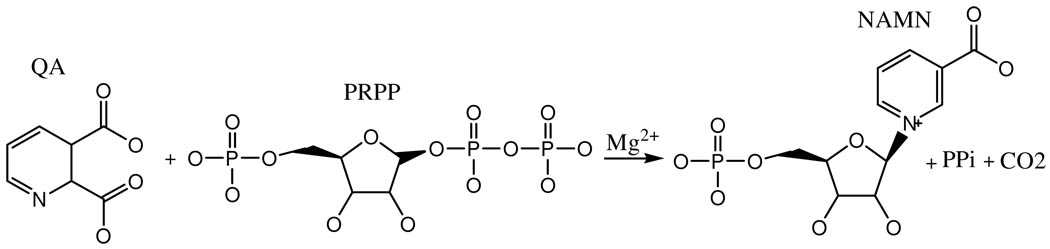
Quinolinate phosphoribosyltransferase (QAPRTase, EC 2.4.2.19) catalyzes an unusual phosphoribosyl transfer that is linked to a decarboxylation reaction to form the NAD precursor nicotinate mononucleotide, carbon dioxide, and pyrophosphate from quinolinic acid (QA) and 5-phosphoribosyl 1-pyrophosphate (PRPP). Structural studies and sequence similarities with other PRTases have implicated Glu214, Asp235, Lys153, and Lys284 in contributing to catalysis through direct interaction with PRPP. The four residues were substituted by site-directed mutagenesis. A nadC deletant form of BL21DE3 was created to eliminate trace contamination by chromosomal QAPRTase. The mutant enzymes were readily purified and retained their dimeric aggregation state on gel filtration. Substitution of Lys153 with Ala resulted in an inactive enzyme, indicating its essential nature. Mutation of Glu214 to Ala or Asp caused at least a 4000-fold reduction in k(cat), with 10-fold increases in K(m) and K(D) values for PRPP. However, mutation of Glu214 to Gln had only modest effects on ligand binding and catalysis. pH profiles indicated that the deprotonated form of a residue with pK(a) of 6.9 is essential for catalysis. The WT-like pH profile of the E214Q mutant indicated that Glu214 is not that residue. Mutation of Asp235 to Ala did not affect ligand binding or catalysis. Mutation of Lys284 to Ala decreased k(cat) by 30-fold and increased K(m) and K(D) values for PRPP by 80-fold and at least 20-fold, respectively. The study suggests that Lys153 is necessary for catalysis and important for PRPP binding, Glu214 provides a hydrogen bond necessary for catalysis but does not act as a base or electrostatically to stabilize the transition state, Lys284 is involved in PRPP binding, and Asp235 is not essential.
Figures

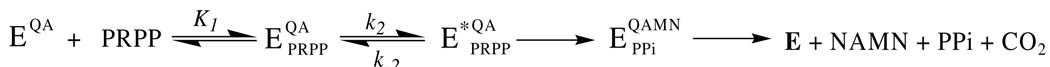
Similar articles
-
Roles for cationic residues at the quinolinic acid binding site of quinolinate phosphoribosyltransferase.Biochemistry. 2010 Feb 23;49(7):1388-95. doi: 10.1021/bi9018225. Biochemistry. 2010. PMID: 20047306 Free PMC article.
-
Structural and kinetic characterization of quinolinate phosphoribosyltransferase (hQPRTase) from homo sapiens.J Mol Biol. 2007 Oct 26;373(3):755-63. doi: 10.1016/j.jmb.2007.08.043. Epub 2007 Aug 24. J Mol Biol. 2007. PMID: 17868694 Free PMC article.
-
Quinolinate phosphoribosyltransferase: kinetic mechanism for a type II PRTase.Biochemistry. 2002 Mar 12;41(10):3520-8. doi: 10.1021/bi012148g. Biochemistry. 2002. PMID: 11876660
-
Role of the flexible loop of hypoxanthine-guanine-xanthine phosphoribosyltransferase from Tritrichomonas foetus in enzyme catalysis.Biochemistry. 2001 Apr 10;40(14):4303-11. doi: 10.1021/bi0026932. Biochemistry. 2001. PMID: 11284686
-
Phosphoribosyl Diphosphate (PRPP): Biosynthesis, Enzymology, Utilization, and Metabolic Significance.Microbiol Mol Biol Rev. 2016 Dec 28;81(1):e00040-16. doi: 10.1128/MMBR.00040-16. Print 2017 Mar. Microbiol Mol Biol Rev. 2016. PMID: 28031352 Free PMC article. Review.
Cited by
-
Roles for cationic residues at the quinolinic acid binding site of quinolinate phosphoribosyltransferase.Biochemistry. 2010 Feb 23;49(7):1388-95. doi: 10.1021/bi9018225. Biochemistry. 2010. PMID: 20047306 Free PMC article.
-
Biochemical characterization of quinolinic acid phosphoribosyltransferase from Mycobacterium tuberculosis H37Rv and inhibition of its activity by pyrazinamide.PLoS One. 2014 Jun 20;9(6):e100062. doi: 10.1371/journal.pone.0100062. eCollection 2014. PLoS One. 2014. PMID: 24949952 Free PMC article.
References
-
- Kurnasov O, Goral V, Colabroy K, Gerdes S, Anantha S, Osterman A, Begley TP. NAD biosynthesis: identification of the tryptophan to quinolinate pathway in bacteria. Chem. Biol. 2003;10:1195–1204. - PubMed
-
- Sharma V, Grubmeyer C, Sacchettini JC. Crystal structure of quinolinic acid phosphoribosyltransferase from Mycobacterium tuberculosis: a potential TB drug target. Structure. 1998;6:1587–1599. - PubMed
-
- Boshoff HI, Xu X, Tahlan K, Dowd CS, Pethe K, Camacho LR, Park T, Yun C, Schnappinger D, Ehrt S, Williams KJ, Barry CE., III Biosynthesis and recycling of nicotinamide cofactors in Mycobacterium tuberculosis. An essential role for NAD in nonreplicating bacilli. J. Biol. Chem. 2008;283:19329–19341. - PMC - PubMed
-
- Schwarcz R, Whetsell WO, Jr, Mangano RM. Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science. 1983;219:316–318. - PubMed
-
- Musick WDL. Structural features of the phosphoribosyltransferases and their relationship to the human deficiency disorders of purine and pyrimidine metabolism. CRC Crit. Rev. Biochem. 1981;11:1–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
