

The evolution and loss of oil-offering flowers: new insights from dated phylogenies for angiosperms and bees
- PMID: 20047869
- PMCID: PMC2838259
- DOI: 10.1098/rstb.2009.0229
The evolution and loss of oil-offering flowers: new insights from dated phylogenies for angiosperms and bees
Abstract
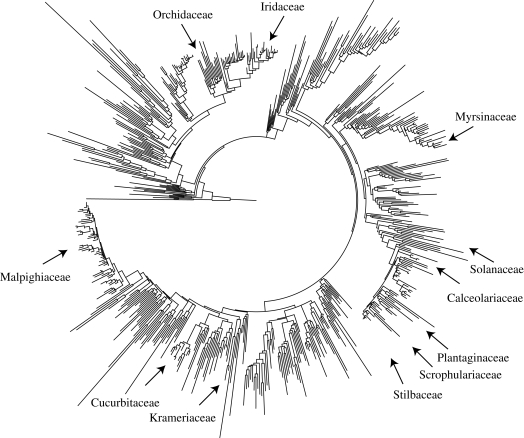
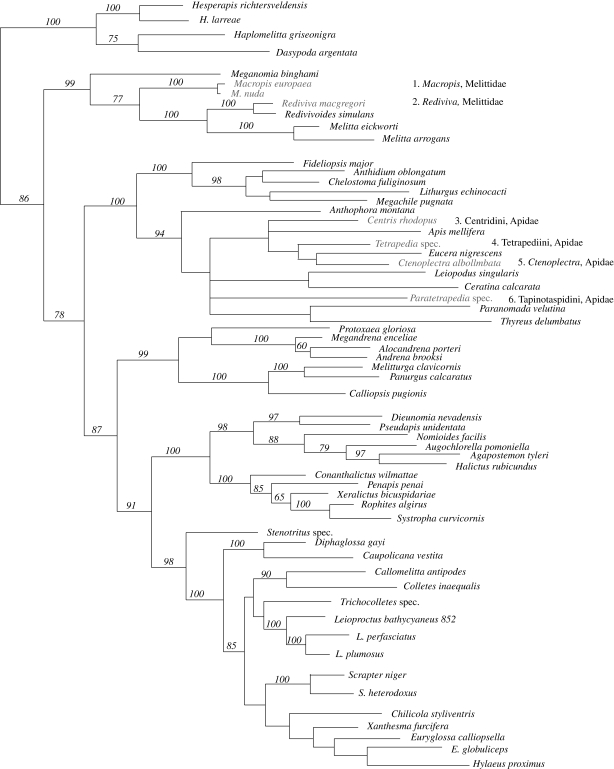
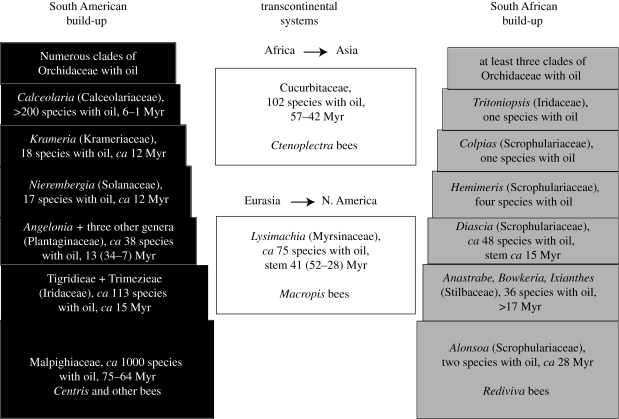
The interactions between bees that depend on floral oil for their larvae and flowers that offer oil involve an intricate mix of obligate and facultative mutualisms. Using recent phylogenies, new data on oil-offering Cucurbitaceae, and molecular-dating, we ask when and how often oil-offering flowers and oil-foraging bees evolved, and how frequently these traits were lost in the cause of evolution. Local phylogenies and an angiosperm-wide tree show that oil flowers evolved at least 28 times and that floral oil was lost at least 36-40 times. The oldest oil flower systems evolved shortly after the K/T boundary independently in American Malpighiaceae, tropical African Cucurbitaceae and Laurasian Lysimachia (Myrsinaceae); the ages of the South African oil flower/oil bee systems are less clear. Youngest oil flower clades include Calceolaria (Calceolariaceae), Iridaceae, Krameria (Krameriaceae) and numerous Orchidaceae, many just a few million years old. In bees, oil foraging evolved minimally seven times and dates back to at least 56 Ma (Ctenoplectra) and 53 Ma (Macropis). The co-occurrence of older and younger oil-offering clades in three of the four geographical regions (but not the Holarctic) implies that oil-foraging bees acquired additional oil hosts over evolutionary time. Such niche-broadening probably started with exploratory visits to flowers resembling oil hosts in scent or colour, as suggested by several cases of Muellerian or Batesian mimicry involving oil flowers.
Figures
Similar articles
-
Flower scent of floral oil-producing Lysimachia punctata as attractant for the oil-bee Macropis fulvipes.J Chem Ecol. 2007 Feb;33(2):441-5. doi: 10.1007/s10886-006-9237-2. Epub 2006 Dec 7. J Chem Ecol. 2007. PMID: 17151908
-
Pterandra pyroidea: a case of pollination shift within neotropical Malpighiaceae.Ann Bot. 2011 Jun;107(8):1323-34. doi: 10.1093/aob/mcr084. Ann Bot. 2011. PMID: 21610210 Free PMC article.
-
Organ-specific volatiles from Sonoran desert Krameria flowers as potential signals for oil-collecting bees.Phytochemistry. 2024 Feb;218:113937. doi: 10.1016/j.phytochem.2023.113937. Epub 2023 Nov 28. Phytochemistry. 2024. PMID: 38035972
-
Time is honey: circadian clocks of bees and flowers and how their interactions may influence ecological communities.Philos Trans R Soc Lond B Biol Sci. 2017 Nov 19;372(1734):20160256. doi: 10.1098/rstb.2016.0256. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28993499 Free PMC article. Review.
-
Buzz pollination: studying bee vibrations on flowers.New Phytol. 2019 Nov;224(3):1068-1074. doi: 10.1111/nph.15666. Epub 2019 Jan 23. New Phytol. 2019. PMID: 30585638 Review.
Cited by
-
Floral rewards in the tribe Sisyrinchieae (Iridaceae): oil as an alternative to pollen and nectar?Sex Plant Reprod. 2012 Dec;25(4):267-79. doi: 10.1007/s00497-012-0196-1. Epub 2012 Sep 12. Sex Plant Reprod. 2012. PMID: 22968405
-
Within and Between-Leg Oil Transfer in an Oil Bee.Integr Org Biol. 2025 Jun 11;7(1):obaf025. doi: 10.1093/iob/obaf025. eCollection 2025. Integr Org Biol. 2025. PMID: 40606674 Free PMC article.
-
Reconstructing an historical pollination syndrome: keel flowers.BMC Ecol Evol. 2022 Apr 12;22(1):45. doi: 10.1186/s12862-022-02003-y. BMC Ecol Evol. 2022. PMID: 35413792 Free PMC article.
-
Insect-flower interactions, ecosystem functions, and restoration ecology in the northern Sahel: current knowledge and perspectives.Biol Rev Camb Philos Soc. 2025 Apr;100(2):969-995. doi: 10.1111/brv.13170. Epub 2024 Nov 26. Biol Rev Camb Philos Soc. 2025. PMID: 39592413 Free PMC article. Review.
-
The temporal build-up of hummingbird/plant mutualisms in North America and temperate South America.BMC Evol Biol. 2015 Jun 10;15:104. doi: 10.1186/s12862-015-0388-z. BMC Evol Biol. 2015. PMID: 26058608 Free PMC article.
References
-
- Aguiar J. C. A., Melo G. A. R.2009Notes on oil sources for the bee genus Caenonomada (Hymenoptera, Apidae, Tapinotaspidini). Rev. Bras. Entomol. 53, 154–156
-
- Aliscioni S. S., Torretta J. P., Bello M. E., Galati G. B.2009Elaiophores in Gomesa bifolia (Sims) M.W. Chase & N.H. Williams (Oncidiinae: Cymbidieae: Orchidaceae): structure and oil secretion. Ann. Bot. 104, 1141–1149 (doi:10.1093/aob/mcp199) - DOI - PMC - PubMed
-
- Alves dos Santos I., Melo G. A. R., Rozen G. R., Jr2002Biology and immature stages of the bee tribe Tetrapediini (Hymenoptera: Apidae). Am. Museum Nov. 3377, 1–45 (doi:10.1206/0003-0082(2002)377<0001:BAISOT>2.0.CO;2) - DOI
-
- Anderberg A. A., Manns U., Källersjö M.2007Phylogeny and floral evolution of the Lysimachieae (Ericales, Myrsinaceae): evidence from ndhF sequence data. Willdenowia 37, 407–421 (doi:10.3372/wi.37.37202) - DOI
-
- Anderson W. R.1979Floral conservatism in Neotropical Malpighiaceae. Biotropica 11, 219–223 (doi:10.2307/2388042) - DOI
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
