

Floral adaptation and diversification under pollen limitation
- PMID: 20047878
- PMCID: PMC2838256
- DOI: 10.1098/rstb.2009.0226
Floral adaptation and diversification under pollen limitation
Abstract
Pollen limitation (PL) of seed production creates unique conditions for reproductive adaptation by angiosperms, in part because, unlike under ovule or resource limitation, floral interactions with pollen vectors can contribute to variation in female success. Although the ecological and conservation consequences of PL have received considerable attention in recent times, its evolutionary implications are poorly appreciated. To identify general influences of PL on reproductive adaptation compared with those under other seed-production limits and their implications for evolution in altered environments, we derive a model that incorporates pollination and post-pollination aspects of PL. Because PL always favours increased ovule fertilization, even when population dynamics are not seed limited, it should pervasively influence selection on reproductive traits. Significantly, under PL the intensity of inbreeding does not determine whether outcrossing or autonomous selfing can evolve, although it can affect which response is most likely. Because the causes of PL are multifaceted in both natural and anthropogenically altered environments, the possible outcrossing solutions are diverse and context dependent, which may contribute to the extensive variety of angiosperm reproductive characteristics. Finally, the increased adaptive options available under PL may be responsible for positive global associations between it and angiosperm diversity.
Figures
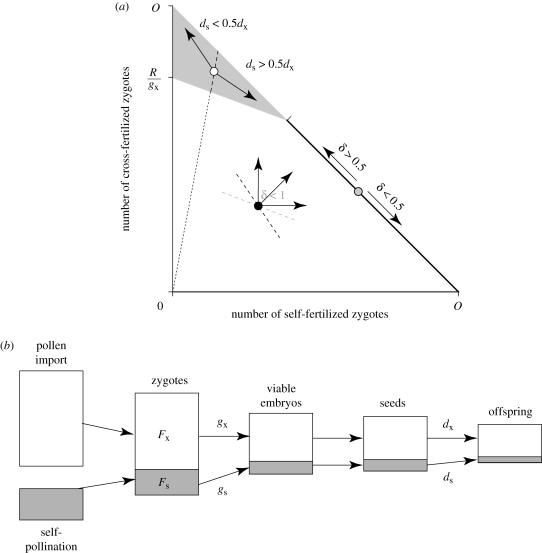
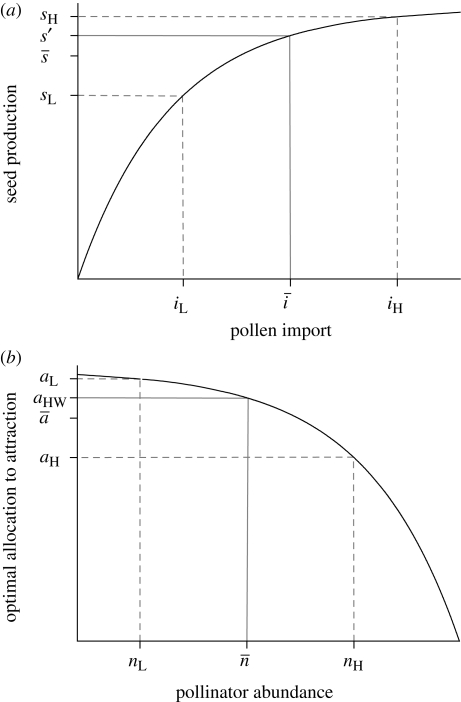
 ) is less than the fecundity expected given the mean pollen import (s′), both of which are lower than the maximum possible seed production, indicating PL. Richards et al. (2009) referred to the difference between s′ and
) is less than the fecundity expected given the mean pollen import (s′), both of which are lower than the maximum possible seed production, indicating PL. Richards et al. (2009) referred to the difference between s′ and  as variance limitation. Similarly, because the optimal allocation to pollinator attraction declines nonlinearly with pollinator abundance, the average optimal allocation to attraction (
as variance limitation. Similarly, because the optimal allocation to pollinator attraction declines nonlinearly with pollinator abundance, the average optimal allocation to attraction ( ) is less than the optimum expected for average pollinator abundance (aHW), which is equivalent to the balance between pollen and resource limitation predicted by Haig & Westoby (1988), resulting in chronic PL.
) is less than the optimum expected for average pollinator abundance (aHW), which is equivalent to the balance between pollen and resource limitation predicted by Haig & Westoby (1988), resulting in chronic PL.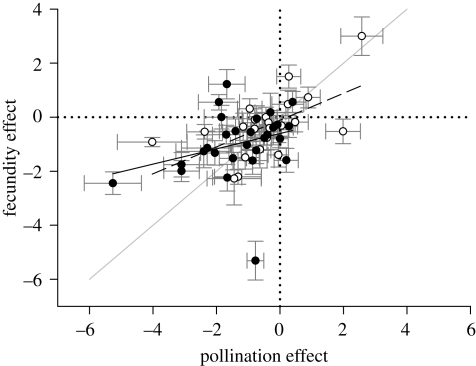
Similar articles
-
Darwin's legacy: the forms, function and sexual diversity of flowers.Philos Trans R Soc Lond B Biol Sci. 2010 Feb 12;365(1539):351-68. doi: 10.1098/rstb.2009.0212. Philos Trans R Soc Lond B Biol Sci. 2010. PMID: 20047864 Free PMC article. Review.
-
Floral longevity and autonomous selfing are altered by pollination and water availability in Collinsia heterophylla.Ann Bot. 2013 Sep;112(5):821-8. doi: 10.1093/aob/mct146. Epub 2013 Jul 24. Ann Bot. 2013. PMID: 23884393 Free PMC article.
-
The Effect of Pollen Limitation on the Evolution of Mating System and Seed Size in Hermaphroditic Plants.Am Nat. 2019 Mar;193(3):447-457. doi: 10.1086/701782. Epub 2019 Jan 18. Am Nat. 2019. PMID: 30794451
-
Variation in floral longevity in the genus Leptosiphon: mating system consequences.Plant Biol (Stuttg). 2013 Jan;15(1):220-5. doi: 10.1111/j.1438-8677.2012.00595.x. Epub 2012 May 18. Plant Biol (Stuttg). 2013. PMID: 22607097
-
Flowering plants under global pollinator decline.Trends Plant Sci. 2013 Jul;18(7):353-9. doi: 10.1016/j.tplants.2013.04.002. Epub 2013 May 18. Trends Plant Sci. 2013. PMID: 23688727 Review.
Cited by
-
GloPL, a global data base on pollen limitation of plant reproduction.Sci Data. 2018 Nov 20;5:180249. doi: 10.1038/sdata.2018.249. Sci Data. 2018. PMID: 30457567 Free PMC article.
-
Pollination and late-acting self-incompatibility in Cyrtanthus breviflorus (Amaryllidaceae): implications for seed production.Ann Bot. 2010 Oct;106(4):547-55. doi: 10.1093/aob/mcq149. Epub 2010 Jul 19. Ann Bot. 2010. PMID: 20647225 Free PMC article.
-
Extremely high proportions of male flowers and geographic variation in floral ratios within male figs of Ficus tikoua despite pollinators displaying active pollen collection.Ecol Evol. 2016 Jan 9;6(2):607-19. doi: 10.1002/ece3.1926. eCollection 2016 Jan. Ecol Evol. 2016. PMID: 26843943 Free PMC article.
-
Adaptation to fragmentation: evolutionary dynamics driven by human influences.Philos Trans R Soc Lond B Biol Sci. 2017 Jan 19;372(1712):20160037. doi: 10.1098/rstb.2016.0037. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 27920382 Free PMC article. Review.
-
Darwin's legacy: the forms, function and sexual diversity of flowers.Philos Trans R Soc Lond B Biol Sci. 2010 Feb 12;365(1539):351-68. doi: 10.1098/rstb.2009.0212. Philos Trans R Soc Lond B Biol Sci. 2010. PMID: 20047864 Free PMC article. Review.
References
-
- Aguilar R., Ashworth L., Galetto L., Aizen M. A.2006Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecol. Lett. 9, 968–980 (doi:10.1111/j.1461-0248.2006.00927.x) - DOI - PubMed
-
- Aizen M. A.2001Flower sex ratio, pollinator abundance, and the seasonal pollination dynamics of a protandrous plant. Ecology 82, 127–144 (doi:10.1890/0012-9658(2001)082[0127:FSRPAA]2.0.CO;2) - DOI
-
- Aizen M. A.2003Down-facing flowers, hummingbirds and rain. Taxon 52, 675–680 (doi:10.2307/3647342) - DOI
-
- Aizen M. A., Feinsinger P.2003Bees not to be? Responses of insect pollinator faunas and flower pollination to habitat fragmentation. In How landscapes change: human disturbance and ecosystem disruptions in the Americas (eds Bradshaw G. A., Marquet P. A.), pp. 111–129 Berlin, Germany: Springer-Verlag
-
- Aizen M. A., Harder L. D.2007Expanding the limits of the pollen-limitation concept: effects of pollen quantity and quality. Ecology 88, 271–281 (doi:10.1890/06-1017) - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
