

A novel mechanism for the scission of double-stranded DNA: BfiI cuts both 3'-5' and 5'-3' strands by rotating a single active site
- PMID: 20047964
- PMCID: PMC2853115
- DOI: 10.1093/nar/gkp1194
A novel mechanism for the scission of double-stranded DNA: BfiI cuts both 3'-5' and 5'-3' strands by rotating a single active site
Abstract
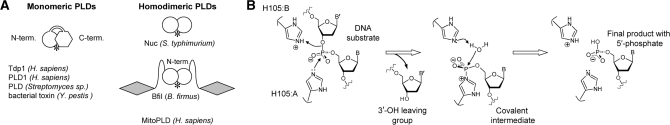
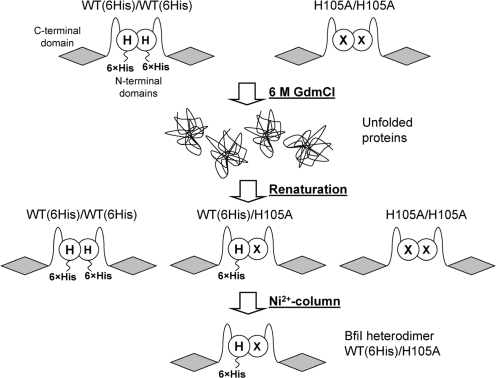
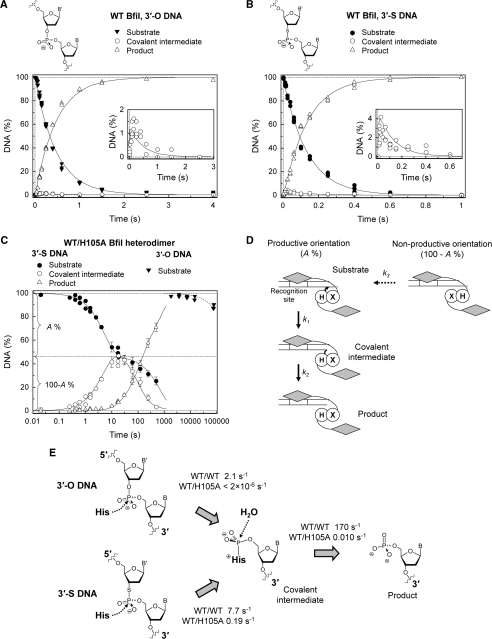
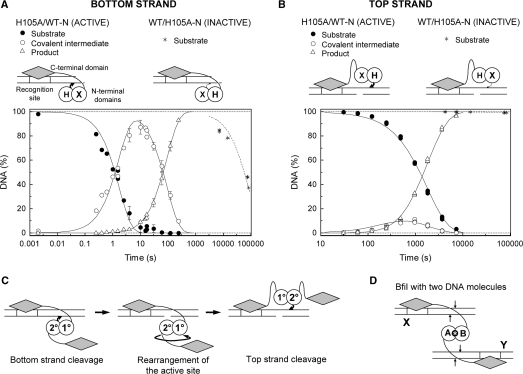
Metal-dependent nucleases that generate double-strand breaks in DNA often possess two symmetrically-equivalent subunits, arranged so that the active sites from each subunit act on opposite DNA strands. Restriction endonuclease BfiI belongs to the phospholipase D (PLD) superfamily and does not require metal ions for DNA cleavage. It exists as a dimer but has at its subunit interface a single active site that acts sequentially on both DNA strands. The active site contains two identical histidines related by 2-fold symmetry, one from each subunit. This symmetrical arrangement raises two questions: first, what is the role and the contribution to catalysis of each His residue; secondly, how does a nuclease with a single active site cut two DNA strands of opposite polarities to generate a double-strand break. In this study, the roles of active-site histidines in catalysis were dissected by analysing heterodimeric variants of BfiI lacking the histidine in one subunit. These variants revealed a novel mechanism for the scission of double-stranded DNA, one that requires a single active site to not only switch between strands but also to switch its orientation on the DNA.
Figures
Similar articles
-
How the BfiI restriction enzyme uses one active site to cut two DNA strands.Proc Natl Acad Sci U S A. 2003 May 27;100(11):6410-5. doi: 10.1073/pnas.1131003100. Epub 2003 May 15. Proc Natl Acad Sci U S A. 2003. PMID: 12750473 Free PMC article.
-
The metal-independent type IIs restriction enzyme BfiI is a dimer that binds two DNA sites but has only one catalytic centre.J Mol Biol. 2003 Feb 28;326(4):1051-64. doi: 10.1016/s0022-2836(03)00020-2. J Mol Biol. 2003. PMID: 12589753
-
Generation of the BfiI restriction endonuclease from the fusion of a DNA recognition domain to a non-specific nuclease from the phospholipase D superfamily.J Mol Biol. 2004 Feb 6;336(1):81-92. doi: 10.1016/j.jmb.2003.12.012. J Mol Biol. 2004. PMID: 14741205
-
Natural and engineered nicking endonucleases--from cleavage mechanism to engineering of strand-specificity.Nucleic Acids Res. 2011 Jan;39(1):1-18. doi: 10.1093/nar/gkq742. Epub 2010 Aug 30. Nucleic Acids Res. 2011. PMID: 20805246 Free PMC article. Review.
-
Artificial restriction DNA cutters as new tools for gene manipulation.Chembiochem. 2009 May 25;10(8):1279-88. doi: 10.1002/cbic.200900040. Chembiochem. 2009. PMID: 19396851 Review.
Cited by
-
Unusual target site disruption by the rare-cutting HNH restriction endonuclease PacI.Structure. 2010 Jun 9;18(6):734-43. doi: 10.1016/j.str.2010.03.009. Structure. 2010. PMID: 20541511 Free PMC article.
-
Catalytically active prokaryotic Argonautes employ phospholipase D family proteins to strengthen immunity against different genetic invaders.mLife. 2024 Sep 4;3(3):403-416. doi: 10.1002/mlf2.12138. eCollection 2024 Sep. mLife. 2024. PMID: 39359674 Free PMC article.
-
DNA binding proteins explore multiple local configurations during docking via rapid rebinding.Nucleic Acids Res. 2016 Sep 30;44(17):8376-84. doi: 10.1093/nar/gkw666. Epub 2016 Jul 28. Nucleic Acids Res. 2016. PMID: 27471033 Free PMC article.
-
Organization of the BcgI restriction-modification protein for the cleavage of eight phosphodiester bonds in DNA.Nucleic Acids Res. 2013 Jan 7;41(1):391-404. doi: 10.1093/nar/gks1023. Epub 2012 Nov 11. Nucleic Acids Res. 2013. PMID: 23147005 Free PMC article.
-
Illuminating the reaction pathway of the FokI restriction endonuclease by fluorescence resonance energy transfer.Nucleic Acids Res. 2012 Feb;40(3):1203-13. doi: 10.1093/nar/gkr809. Epub 2011 Oct 12. Nucleic Acids Res. 2012. PMID: 21993298 Free PMC article.
References
-
- Horton NC. In: Protein-Nucleic Acid Interactions: Structural Biology. Rice PA, Correll CC, editors. Cambridge, UK: RSC Publishing; 2008.
-
- Connolly BA, Eckstein F, Pingoud A. The stereochemical course of the restriction endonuclease EcoRI-catalyzed reaction. J. Biol. Chem. 1984;259:10760–10763. - PubMed
-
- Elliott SL, Brazier J, Cosstick R, Connolly BA. Mechanism of the I DNA T:G-mismatch endonuclease (Vsr protein) probed with thiophosphate-containing oligodeoxynucleotides. J. Mol. Biol. 2005;353:692–703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
