

Location analysis for the estrogen receptor-alpha reveals binding to diverse ERE sequences and widespread binding within repetitive DNA elements
- PMID: 20047966
- PMCID: PMC2853111
- DOI: 10.1093/nar/gkp1188
Location analysis for the estrogen receptor-alpha reveals binding to diverse ERE sequences and widespread binding within repetitive DNA elements
Abstract
Location analysis for estrogen receptor-alpha (ERalpha)-bound cis-regulatory elements was determined in MCF7 cells using chromatin immunoprecipitation (ChIP)-on-chip. Here, we present the estrogen response element (ERE) sequences that were identified at ERalpha-bound loci and quantify the incidence of ERE sequences under two stringencies of detection: <10% and 10-20% nucleotide deviation from the canonical ERE sequence. We demonstrate that approximately 50% of all ERalpha-bound loci do not have a discernable ERE and show that most ERalpha-bound EREs are not perfect consensus EREs. Approximately one-third of all ERalpha-bound ERE sequences reside within repetitive DNA sequences, most commonly of the AluS family. In addition, the 3-bp spacer between the inverted ERE half-sites, rather than being random nucleotides, is C(A/T)G-enriched at bona fide receptor targets. Diverse ERalpha-bound loci were validated using electrophoretic mobility shift assay and ChIP-polymerase chain reaction (PCR). The functional significance of receptor-bound loci was demonstrated using luciferase reporter assays which proved that repetitive element ERE sequences contribute to enhancer function. ChIP-PCR demonstrated estrogen-dependent recruitment of the coactivator SRC3 to these loci in vivo. Our data demonstrate that ERalpha binds to widely variant EREs with less sequence specificity than had previously been suspected and that binding at repetitive and nonrepetitive genomic targets is favored by specific trinucleotide spacers.
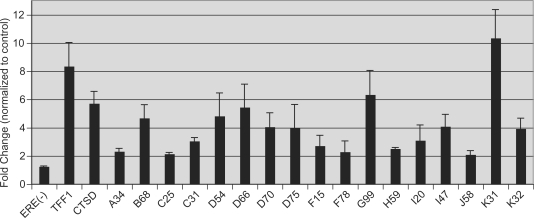
Figures





Similar articles
-
The tri-nucleotide spacer sequence between estrogen response element half-sites is conserved and modulates ERalpha-mediated transcriptional responses.J Steroid Biochem Mol Biol. 2010 Jun;120(4-5):172-9. doi: 10.1016/j.jsbmb.2010.04.009. Epub 2010 Apr 18. J Steroid Biochem Mol Biol. 2010. PMID: 20403436 Free PMC article.
-
Intercalation of XR5944 with the estrogen response element is modulated by the tri-nucleotide spacer sequence between half-sites.J Steroid Biochem Mol Biol. 2011 Apr;124(3-5):121-7. doi: 10.1016/j.jsbmb.2011.02.003. Epub 2011 Feb 17. J Steroid Biochem Mol Biol. 2011. PMID: 21333738 Free PMC article.
-
Role of estrogen receptor ligand and estrogen response element sequence on interaction with chicken ovalbumin upstream promoter transcription factor (COUP-TF).J Steroid Biochem Mol Biol. 1999 Nov;71(1-2):1-19. doi: 10.1016/s0960-0760(99)00124-7. J Steroid Biochem Mol Biol. 1999. PMID: 10619353
-
Estrogen response element-dependent regulation of transcriptional activation of estrogen receptors alpha and beta by coactivators and corepressors.J Mol Endocrinol. 2004 Oct;33(2):387-410. doi: 10.1677/jme.1.01541. J Mol Endocrinol. 2004. PMID: 15525597
-
Two estrogen response element sequences near the PCNA gene are not responsible for its estrogen-enhanced expression in MCF7 cells.PLoS One. 2008;3(10):e3523. doi: 10.1371/journal.pone.0003523. Epub 2008 Oct 24. PLoS One. 2008. PMID: 18949048 Free PMC article.
Cited by
-
BRCA1-mimetic compound NSC35446.HCl inhibits IKKB expression by reducing estrogen receptor-α occupancy in the IKKB promoter and inhibits NF-κB activity in antiestrogen-resistant human breast cancer cells.Breast Cancer Res Treat. 2017 Dec;166(3):681-693. doi: 10.1007/s10549-017-4442-y. Epub 2017 Aug 14. Breast Cancer Res Treat. 2017. PMID: 28808806 Free PMC article.
-
The Regulation and Immune Signature of Retrotransposons in Cancer.Cancers (Basel). 2023 Aug 30;15(17):4340. doi: 10.3390/cancers15174340. Cancers (Basel). 2023. PMID: 37686616 Free PMC article. Review.
-
Phosphoproteome Analysis Reveals Estrogen-ER Pathway as a Modulator of mTOR Activity Via DEPTOR.Mol Cell Proteomics. 2019 Aug;18(8):1607-1618. doi: 10.1074/mcp.RA119.001506. Epub 2019 Jun 12. Mol Cell Proteomics. 2019. PMID: 31189691 Free PMC article.
-
From Influenza Virus Infections to Lupus: Synchronous Estrogen Receptor α and RNA Polymerase II Binding Within the Immunoglobulin Heavy Chain Locus.Viral Immunol. 2020 May;33(4):307-315. doi: 10.1089/vim.2019.0144. Epub 2020 Feb 27. Viral Immunol. 2020. PMID: 32105583 Free PMC article.
-
Global investigation of estrogen-responsive genes regulating lipid metabolism in the liver of laying hens.BMC Genomics. 2021 Jun 9;22(1):428. doi: 10.1186/s12864-021-07679-y. BMC Genomics. 2021. PMID: 34107898 Free PMC article.
References
-
- Yager JD, Davidson NE. Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 2006;354:270–282. - PubMed
-
- Smith CL, O'M;alley BW. Coregulator function: a key to understanding tissue specificity of selective receptor modulators. Endocr. Rev. 2004;25:45–71. - PubMed
-
- Bjornstrom L, Sjoberg M. Mechanisms of estrogen receptor signaling: convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol. 2005;19:833–842. - PubMed
-
- Cerillo G, Rees A, Manchanda N, Reilly C, Brogan I, White A, Needham M. The oestrogen receptor regulates NFkappaB and AP-1 activity in a cell-specific manner. J. Steroid Biochem. Mol. Biol. 1998;67:79–88. - PubMed
-
- Paech K, Webb P, Kuiper GG, Nilsson S, Gustafsson J, Kushner PJ, Scanlan TS. Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sites. Science. 1997;277:1508–1510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
