

NF45 and NF90 regulate HS4-dependent interleukin-13 transcription in T cells
- PMID: 20051514
- PMCID: PMC2832977
- DOI: 10.1074/jbc.M109.041004
NF45 and NF90 regulate HS4-dependent interleukin-13 transcription in T cells
Abstract
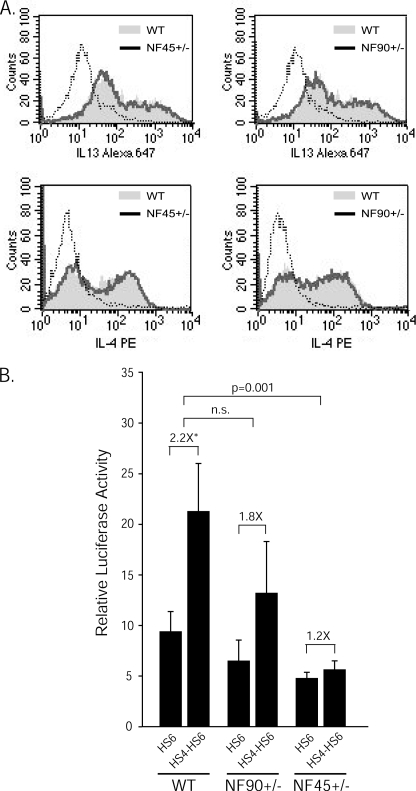
Expression of the cytokine interleukin-13 (IL13) is critical for Th2 immune responses and Th2-mediated allergic diseases. Activation of human IL13 expression involves chromatin remodeling and formation of multiple DNase I-hypersensitive sites throughout the locus. Among these, HS4 is detected in the distal IL13 promoter in both naive and polarized CD4(+) T cells. We show herein that HS4 acts as a position-independent, orientation-dependent positive regulator of IL13 proximal promoter activity in transiently transfected, activated human CD4(+) Jurkat T cells and primary murine Th2 cells. The 3'-half of HS4 (HS4-3') was responsible for IL13 up-regulation and bound nuclear factor (NF) 90 and NF45, as demonstrated by DNA affinity chromatography coupled with tandem mass spectrometry, chromatin immunoprecipitation, and gel shift analysis. Notably, the CTGTT NF45/NF90-binding motif within HS4-3' was critical for HS4-dependent up-regulation of IL13 expression. Moreover, transfection of HS4-IL13 reporter vectors into primary, in vitro differentiated Th2 cells from wild-type, NF45(+/-), or NF90(+/-) mice showed that HS4 activity was exquisitely dependent on the levels of endogenous NF45 (and to a lesser degree NF90), because HS4-dependent IL13 expression was virtually abrogated in NF45(+/-) cells and reduced in NF90(+/-) cells. Collectively, our results identify NF45 and NF90 as novel regulators of HS4-dependent human IL13 transcription in response to T cell activation.
Figures






Similar articles
-
An allergy-associated polymorphism in a novel regulatory element enhances IL13 expression.Hum Mol Genet. 2009 Dec 1;18(23):4513-20. doi: 10.1093/hmg/ddp411. Epub 2009 Aug 25. Hum Mol Genet. 2009. PMID: 19706623 Free PMC article.
-
NF45 and NF90 in murine seminiferous epithelium: potential role in SP-10 gene transcription.J Androl. 2008 Mar-Apr;29(2):186-97. doi: 10.2164/jandrol.107.003756. Epub 2007 Oct 17. J Androl. 2008. PMID: 17942973
-
Dynamic binding of Ku80, Ku70 and NF90 to the IL-2 promoter in vivo in activated T-cells.Nucleic Acids Res. 2007;35(7):2302-10. doi: 10.1093/nar/gkm117. Epub 2007 Mar 27. Nucleic Acids Res. 2007. PMID: 17389650 Free PMC article.
-
Inducible expression of immediate early genes is regulated through dynamic chromatin association by NF45/ILF2 and NF90/NF110/ILF3.PLoS One. 2019 Apr 25;14(4):e0216042. doi: 10.1371/journal.pone.0216042. eCollection 2019. PLoS One. 2019. PMID: 31022259 Free PMC article.
-
NF45/NF90-mediated rDNA transcription provides a novel target for immunosuppressant development.EMBO Mol Med. 2021 Mar 5;13(3):e12834. doi: 10.15252/emmm.202012834. Epub 2021 Feb 8. EMBO Mol Med. 2021. PMID: 33555115 Free PMC article.
Cited by
-
The lncRNA SNHG26 drives the inflammatory-to-proliferative state transition of keratinocyte progenitor cells during wound healing.Nat Commun. 2024 Oct 5;15(1):8637. doi: 10.1038/s41467-024-52783-8. Nat Commun. 2024. PMID: 39366968 Free PMC article.
-
NF90-NF45 is essential for β cell compensation under obesity-inducing metabolic stress through suppression of p53 signaling pathway.Sci Rep. 2022 May 25;12(1):8837. doi: 10.1038/s41598-022-12600-y. Sci Rep. 2022. PMID: 35614067 Free PMC article.
-
Expression and Clinical Significance of ILF2 in Gastric Cancer.Dis Markers. 2017;2017:4387081. doi: 10.1155/2017/4387081. Epub 2017 Jul 31. Dis Markers. 2017. PMID: 28831206 Free PMC article.
-
Network analysis of quantitative proteomics on asthmatic bronchi: effects of inhaled glucocorticoid treatment.Respir Res. 2011 Sep 22;12(1):124. doi: 10.1186/1465-9921-12-124. Respir Res. 2011. PMID: 21939520 Free PMC article. Clinical Trial.
-
The long noncoding RNA LINC00473, a target of microRNA 34a, promotes tumorigenesis by inhibiting ILF2 degradation in cervical cancer.Am J Cancer Res. 2017 Nov 1;7(11):2157-2168. eCollection 2017. Am J Cancer Res. 2017. PMID: 29218240 Free PMC article.
References
-
- Finkelman F. D., Shea-Donohue T., Morris S. C., Gildea L., Strait R., Madden K. B., Schopf L., Urban J. F., Jr. (2004) Immunol. Rev. 201, 139–155 - PubMed
-
- Vercelli D. (2008) Nat. Rev. Immunol. 8, 169–182 - PubMed
-
- Wills-Karp M., Luyimbazi J., Xu X., Schofield B., Neben T. Y., Karp C. L., Donaldson D. D. (1998) Science 282, 2258–2261 - PubMed
-
- Walter D. M., McIntire J. J., Berry G., McKenzie A. N., Donaldson D. D., DeKruyff R. H., Umetsu D. T. (2001) J. Immunol. 167, 4668–4675 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
