

HSF1-controlled and age-associated chaperone capacity in neurons and muscle cells of C. elegans
- PMID: 20052290
- PMCID: PMC2797298
- DOI: 10.1371/journal.pone.0008568
HSF1-controlled and age-associated chaperone capacity in neurons and muscle cells of C. elegans
Abstract
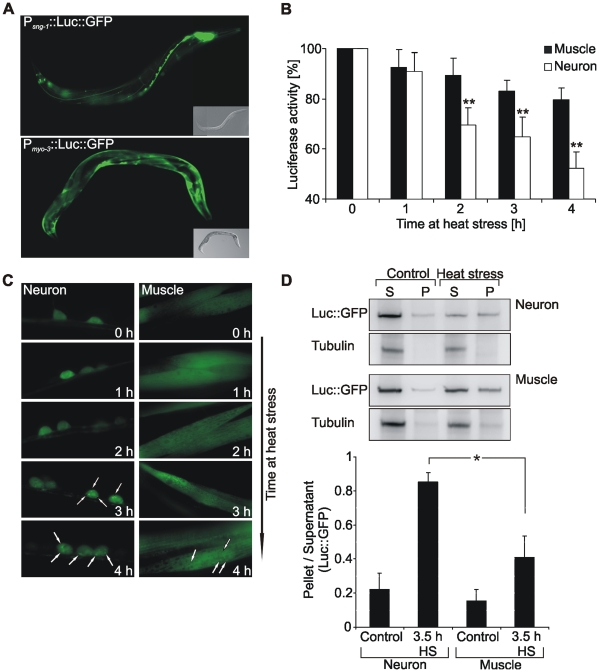
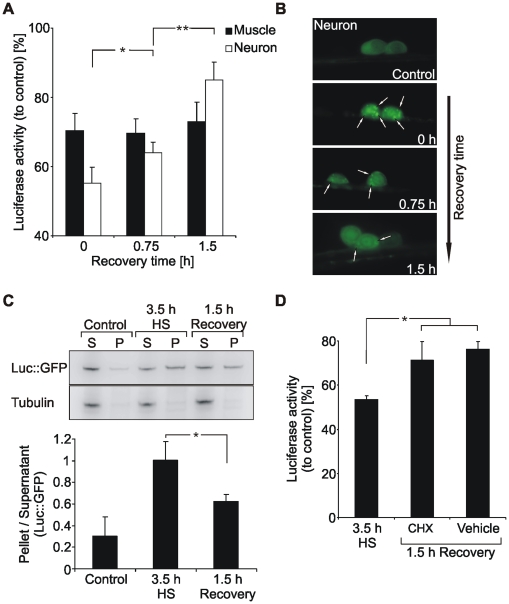
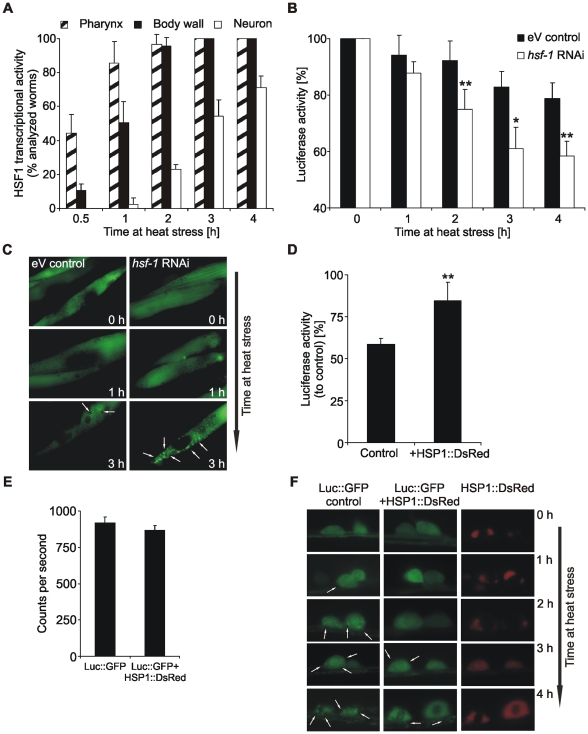
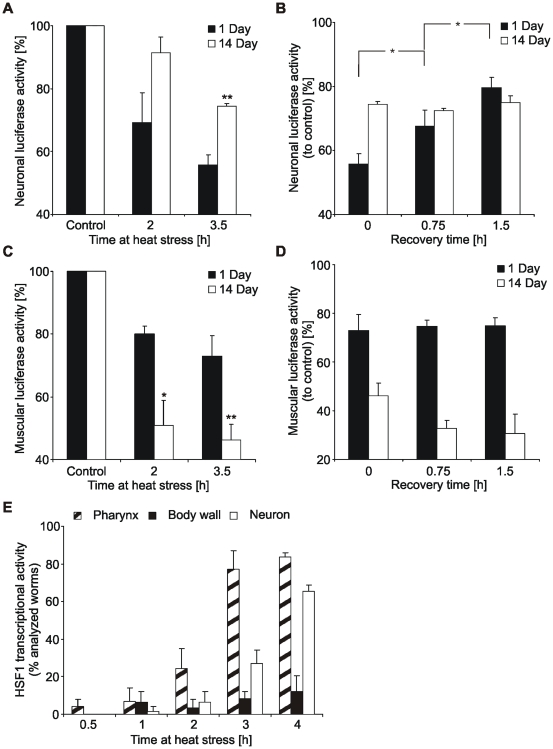
Protein stability under changing conditions is of vital importance for the cell and under the control of a fine-tuned network of molecular chaperones. Aging and age-related neurodegenerative diseases are directly associated with enhanced protein instability. Employing C. elegans expressing GFP-tagged luciferase as a reporter for evaluation of protein stability we show that the chaperoning strategy of body wall muscle cells and neurons is significantly different and that both are differently affected by aging. Muscle cells of young worms are largely resistant to heat stress, which is directly mediated by the stress response controlled through Heat Shock Transcription Factor 1. During recovery following heat stress the ability to refold misfolded proteins is missing. Young neurons are highly susceptible to chronic heat stress, but show a high potency to refold or disaggregate proteins during subsequent recovery. The particular proteome instability in neurons results from a delayed induction of the heat shock response. In aged neurons protein stability is increased during heat stress, whereas muscle cells show enhanced protein instability due to a deteriorated heat shock response. An efficient refolding activity is absent in both aged tissues. These results provide molecular insights into the differential protein stabilization capacity in different tissues and during aging.
Conflict of interest statement
Figures
Similar articles
-
Lifespan extension in C. elegans by a molecular chaperone dependent upon insulin-like signals.Aging Cell. 2003 Apr;2(2):131-9. doi: 10.1046/j.1474-9728.2003.00045.x. Aging Cell. 2003. PMID: 12882326
-
C. elegans STI-1, the homolog of Sti1/Hop, is involved in aging and stress response.J Mol Biol. 2009 Jul 24;390(4):604-17. doi: 10.1016/j.jmb.2009.05.035. Epub 2009 May 23. J Mol Biol. 2009. PMID: 19467242
-
A neuronal GPCR is critical for the induction of the heat shock response in the nematode C. elegans.J Neurosci. 2013 Apr 3;33(14):6102-11. doi: 10.1523/JNEUROSCI.4023-12.2013. J Neurosci. 2013. PMID: 23554491 Free PMC article.
-
Longevity and heat stress regulation in Caenorhabditis elegans.Mech Ageing Dev. 2003 Jan;124(1):43-8. doi: 10.1016/s0047-6374(02)00168-9. Mech Ageing Dev. 2003. PMID: 12618005 Review.
-
Feedback regulation of the heat shock response.Handb Exp Pharmacol. 2006;(172):43-68. doi: 10.1007/3-540-29717-0_2. Handb Exp Pharmacol. 2006. PMID: 16610354 Review.
Cited by
-
Cloning and Transcriptional Activity of the Mouse Omi/HtrA2 Gene Promoter.Int J Mol Sci. 2016 Jan 16;17(1):119. doi: 10.3390/ijms17010119. Int J Mol Sci. 2016. PMID: 26784188 Free PMC article.
-
Investigating the spreading and toxicity of prion-like proteins using the metazoan model organism C. elegans.J Vis Exp. 2015 Jan 8;(95):52321. doi: 10.3791/52321. J Vis Exp. 2015. PMID: 25591151 Free PMC article.
-
Glucose delays age-dependent proteotoxicity.Aging Cell. 2012 Oct;11(5):856-66. doi: 10.1111/j.1474-9726.2012.00855.x. Epub 2012 Aug 1. Aging Cell. 2012. PMID: 22734670 Free PMC article.
-
Receptor-mediated endocytosis 8 (RME-8)/DNAJC13 is a novel positive modulator of autophagy and stabilizes cellular protein homeostasis.Cell Mol Life Sci. 2021 Jan;78(2):645-660. doi: 10.1007/s00018-020-03521-y. Epub 2020 Apr 22. Cell Mol Life Sci. 2021. PMID: 32322926 Free PMC article.
-
Cellular stress response pathways and ageing: intricate molecular relationships.EMBO J. 2011 May 17;30(13):2520-31. doi: 10.1038/emboj.2011.162. EMBO J. 2011. PMID: 21587205 Free PMC article. Review.
References
-
- Tartaglia GG, Pechmann S, Dobson CM, Vendruscolo M. Life on the edge: a link between gene expression levels and aggregation rates of human proteins. Trends Biochem Sci. 2007;32:204–206. - PubMed
-
- Zimmerman SB, Minton AP. Macromolecular crowding: biochemical, biophysical, and physiological consequences. Annu Rev Biophys Biomol Struct. 1993;22:27–65. - PubMed
-
- Chang HC, Tang YC, Hayer-Hartl M, Hartl FU. SnapShot: molecular chaperones, Part I. Cell. 2007;128:212. - PubMed
-
- Hartl FU, Hayer-Hartl M. Molecular chaperones in the cytosol: from nascent chain to folded protein. Science. 2002;295:1852–1858. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
