

Recombinant prion protein induces a new transmissible prion disease in wild-type animals
- PMID: 20052481
- PMCID: PMC2808531
- DOI: 10.1007/s00401-009-0633-x
Recombinant prion protein induces a new transmissible prion disease in wild-type animals
Abstract
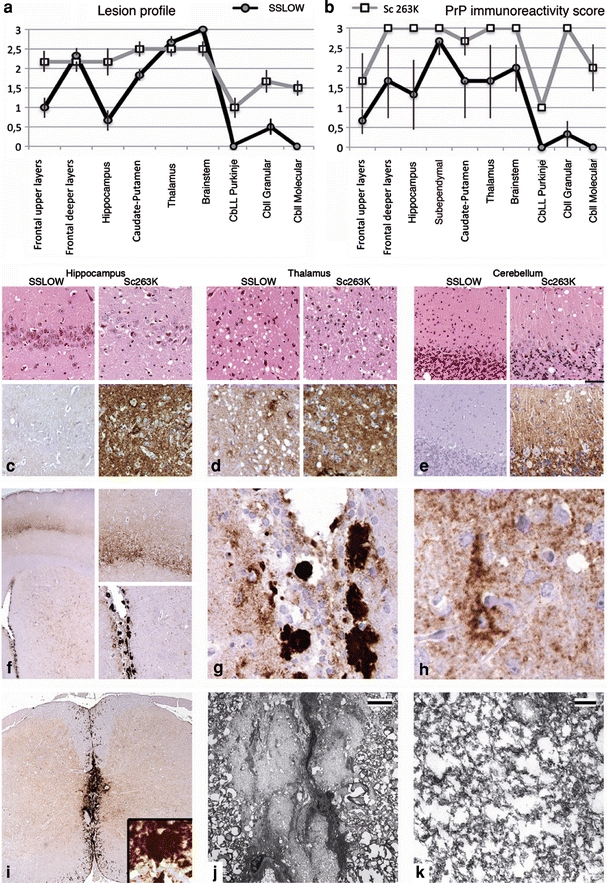
Prion disease is a neurodegenerative malady, which is believed to be transmitted via a prion protein in its abnormal conformation (PrP(Sc)). Previous studies have failed to demonstrate that prion disease could be induced in wild-type animals using recombinant prion protein (rPrP) produced in Escherichia coli. Here, we report that prion infectivity was generated in Syrian hamsters after inoculating full-length rPrP that had been converted into the cross-beta-sheet amyloid form and subjected to annealing. Serial transmission gave rise to a disease phenotype with highly unique clinical and neuropathological features. Among them were the deposition of large PrP(Sc) plaques in subpial and subependymal areas in brain and spinal cord, very minor lesioning of the hippocampus and cerebellum, and a very slow progression of disease after onset of clinical signs despite the accumulation of large amounts of PrP(Sc) in the brain. The length of the clinical duration is more typical of human and large animal prion diseases, than those of rodents. Our studies establish that transmissible prion disease can be induced in wild-type animals by inoculation of rPrP and introduce a valuable new model of prion diseases.
Figures
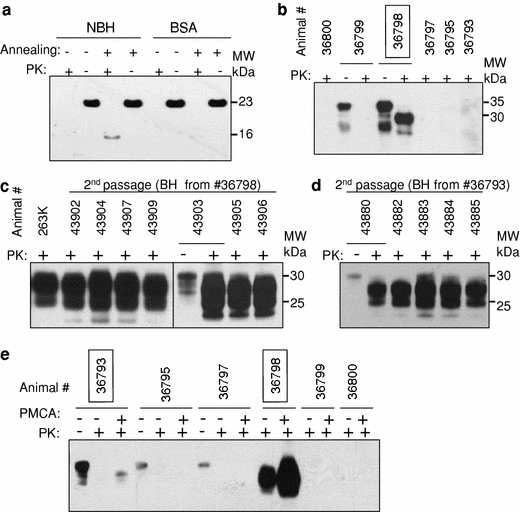
References
-
- Baskakov IV, Breydo L. Converting the prion protein: what makes the protein infectious. Biochim Biophys Acta (Molecular Basis of Disease) 2007;1772:692–703. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
