

Two kinetic patterns of epitope-specific CD8 T-cell responses following murine gammaherpesvirus 68 infection
- PMID: 20053740
- PMCID: PMC2826075
- DOI: 10.1128/JVI.02229-09
Two kinetic patterns of epitope-specific CD8 T-cell responses following murine gammaherpesvirus 68 infection
Abstract
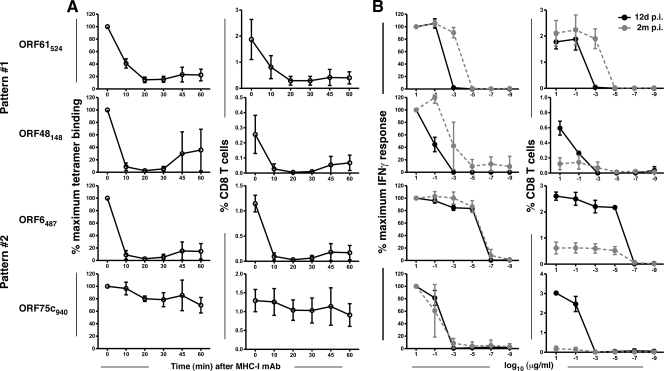
Murine gammaherpesvirus 68 (gammaHV68) provides an important experimental model for understanding mechanisms of immune control of the latent human gammaherpesviruses. Antiviral CD8 T cells play a key role throughout three separate phases of the infection: clearance of lytic virus, control of the latency amplification stage, and prevention of reactivation of latently infected cells. Previous analyses have shown that T-cell responses to two well-characterized epitopes derived from ORF6 and ORF61 progress with distinct kinetics. ORF6(487)-specific cells predominate early in infection and then decline rapidly, whereas ORF61(524)-specific cells continue to expand through early latency, due to sustained epitope expression. However, the paucity of identified epitopes to this virus has limited our understanding of the overall complexities of CD8 T-cell immune control throughout infection. Here we screened 1,383 predicted H-2(b)-restricted peptides and identified 33 responses, of which 21 have not previously been reported. Kinetic analysis revealed a spectrum of T-cell responses based on the rapidity of their decline after the peak acute response that generally corresponded to the expression patterns of the two previously characterized epitopes. The slowly declining responses that were maintained during latency amplification proliferated more rapidly and underwent maturation of functional avidity over time. Furthermore, the kinetics of decline was accelerated following infection with a latency-null mutant virus. Overall, the data show that gammaHV68 infection elicits a highly heterogeneous CD8 T-cell response that segregates into two distinctive kinetic patterns controlled by differential epitope expression during the lytic and latency amplification stages of infection.
Figures








References
-
- Alter, G., J. M. Malenfant, and M. Altfeld. 2004. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 294:15-22. - PubMed
-
- Cheng, Y., and W. H. Prusoff. 1973. Relationship between the inhibition constant (K1) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem. Pharmacol. 22:3099-3108. - PubMed
-
- Coppola, M. A., E. Flano, P. Nguyen, C. L. Hardy, R. D. Cardin, N. Shastri, D. L. Woodland, and M. A. Blackman. 1999. Apparent MHC-independent stimulation of CD8+ T cells in vivo during latent murine gammaherpesvirus infection. J. Immunol. 163:1481-1489. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- CA148250/CA/NCI NIH HHS/United States
- R01 DE015752/DE/NIDCR NIH HHS/United States
- R01 DE014153/DE/NIDCR NIH HHS/United States
- DE15752/DE/NIDCR NIH HHS/United States
- F32 AI084327/AI/NIAID NIH HHS/United States
- DE14153/DE/NIDCR NIH HHS/United States
- R01 CA091791/CA/NCI NIH HHS/United States
- RC2 CA148250/CA/NCI NIH HHS/United States
- N01-AI400024C/AI/NIAID NIH HHS/United States
- AI42927/AI/NIAID NIH HHS/United States
- R21 DE018337/DE/NIDCR NIH HHS/United States
- T32 AI049823/AI/NIAID NIH HHS/United States
- DE18337/DE/NIDCR NIH HHS/United States
- T32 AI49823/AI/NIAID NIH HHS/United States
- R01 AI042927/AI/NIAID NIH HHS/United States
- CA91791/CA/NCI NIH HHS/United States
- N01-AI400023C/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
