

Delayed intrinsic activation of an NMDA-independent CaM-kinase II in a critical time window is necessary for late consolidation of an associative memory
- PMID: 20053887
- PMCID: PMC6632524
- DOI: 10.1523/JNEUROSCI.2577-09.2010
Delayed intrinsic activation of an NMDA-independent CaM-kinase II in a critical time window is necessary for late consolidation of an associative memory
Abstract
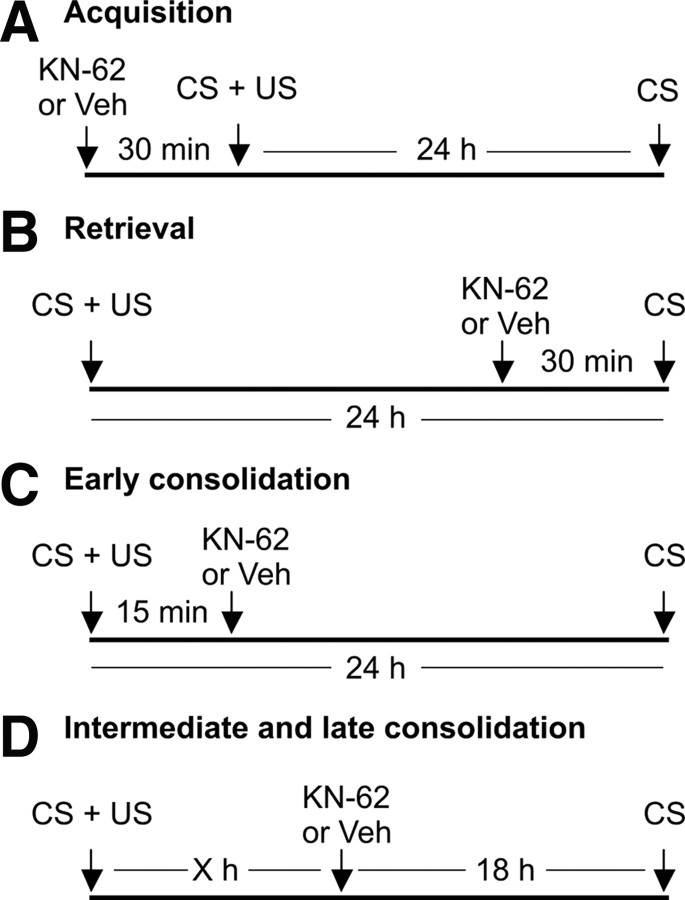
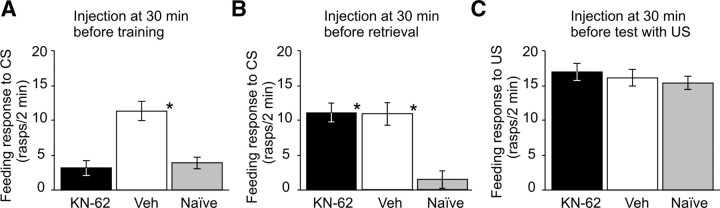
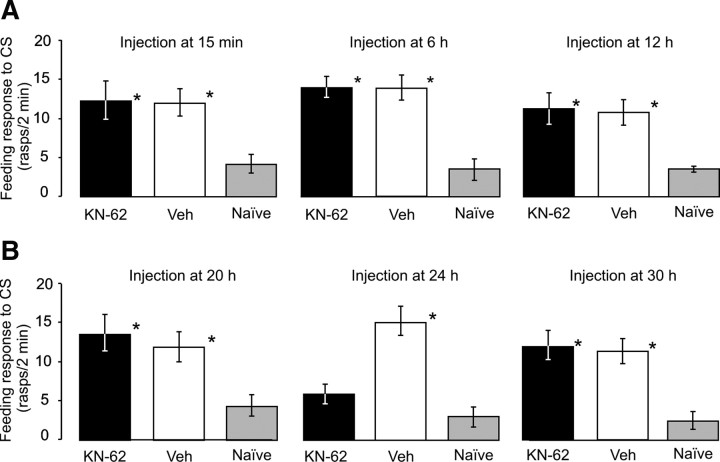
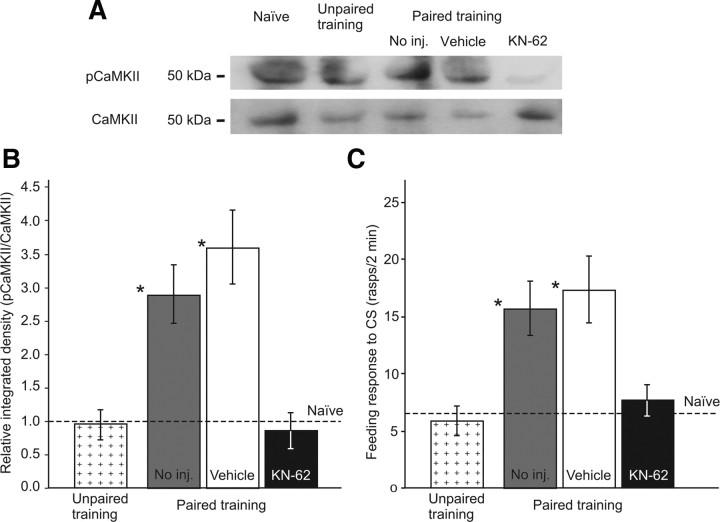
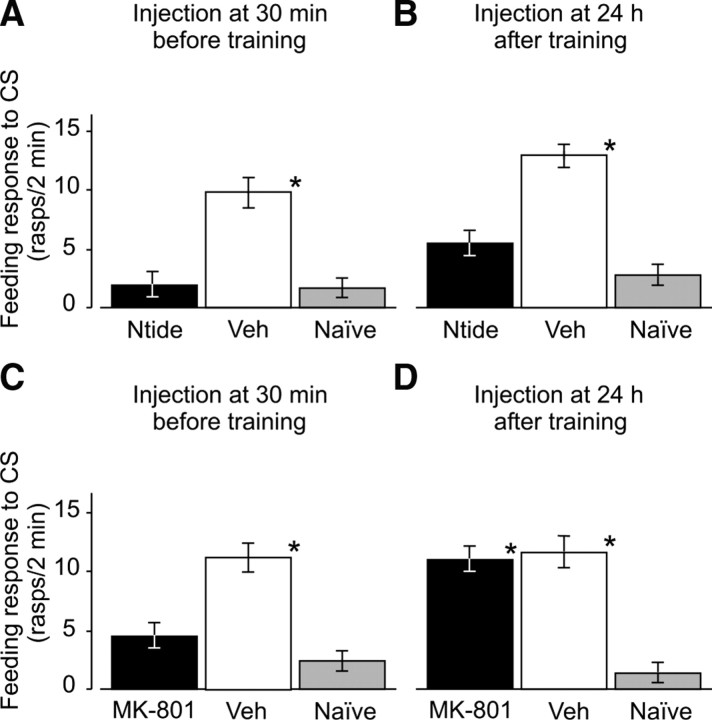
Calcium/calmodulin-dependent kinases (CaM-kinases) are central to various forms of long-term memory (LTM) in a number of evolutionarily diverse organisms. However, it is still largely unknown what contributions specific CaM-kinases make to different phases of the same specific type of memory, such as acquisition, or early, intermediate, and late consolidation of associative LTM after classical conditioning. Here, we investigated the involvement of CaM-kinase II (CaMKII) in different phases of associative LTM induced by single-trial reward classical conditioning in Lymnaea, a well established invertebrate experimental system for studying molecular mechanisms of learning and memory. First, by using a general CaM-kinase inhibitor, KN-62, we found that CaM-kinase activation was necessary for acquisition and late consolidation, but not early or intermediate consolidation or retrieval of LTM. Then, we used Western blot-based phosphorylation assays and treatment with CaMKIINtide to identify CaMKII as the main CaM-kinase, the intrinsic activation of which, in a critical time window ( approximately 24 h after learning), is central to late consolidation of LTM. Additionally, using MK-801 and CaMKIINtide we found that acquisition was dependent on both NMDA receptor and CaMKII activation. However, unlike acquisition, CaMKII-dependent late memory consolidation does not require the activation of NMDA receptors. Our new findings support the notion that even apparently stable memory traces may undergo further molecular changes and identify NMDA-independent intrinsic activation of CaMKII as a mechanism underlying this "lingering consolidation." This process may facilitate the preservation of LTM in the face of protein turnover or active molecular processes that underlie forgetting.
Figures

References
-
- Abel T, Lattal KM. Molecular mechanisms of memory acquisition, consolidation and retrieval. Curr Opin Neurobiol. 2001;11:180–187. - PubMed
-
- Alexander J, Jr, Audesirk TE, Audesirk GJ. One-trial reward learning in the snail Lymnaea stagnalis. J Neurobiol. 1984;15:67–72. - PubMed
-
- Bayer KU, De Koninck P, Leonard AS, Hell JW, Schulman H. Interaction with the NMDA receptor locks CaMKII in an active conformation. Nature. 2001;411:801–805. - PubMed
-
- Bevilaqua LR, Medina JH, Izquierdo I, Cammarota M. Memory consolidation induces N-methyl-D-aspartic acid-receptor- and Ca2+/calmodulin-dependent protein kinase II-dependent modifications in α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor properties. Neuroscience. 2005;136:397–403. - PubMed
-
- Brierley MJ, Yeoman MS, Benjamin PR. Glutamate is the transmitter for N2v retraction phase interneurons of the Lymnaea feeding system. J Neurophysiol. 1997;78:3408–3414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical