

Functional role of gamma and theta oscillations in episodic memory
- PMID: 20060015
- PMCID: PMC2856712
- DOI: 10.1016/j.neubiorev.2009.12.014
Functional role of gamma and theta oscillations in episodic memory
Abstract
The primary aim of this review is to examine evidence for a functional role of gamma and theta oscillations in human episodic memory. It is proposed here that gamma and theta oscillations allow for the transient interaction between cortical structures and the hippocampus for the encoding and retrieval of episodic memories as described by the hippocampal memory indexing theory (Teyler and DiScenna, 1986). Gamma rhythms can act in the cortex to bind perceptual features and in the hippocampus to bind the rich perceptual and contextual information from diverse brain regions into episodic representations. Theta oscillations act to temporally order these individual episodic memory representations. Through feedback projections from the hippocampus to the cortex these gamma and theta patterns could cause the reinstatement of the entire episodic memory representation in the cortex. In addition, theta oscillations could allow for top-down control from the frontal cortex to the hippocampus modulating the encoding and retrieval of episodic memories.
Figures
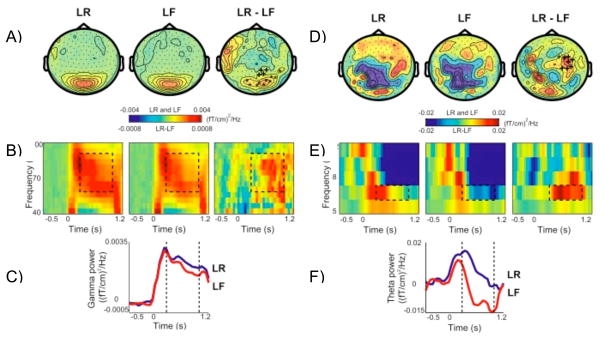
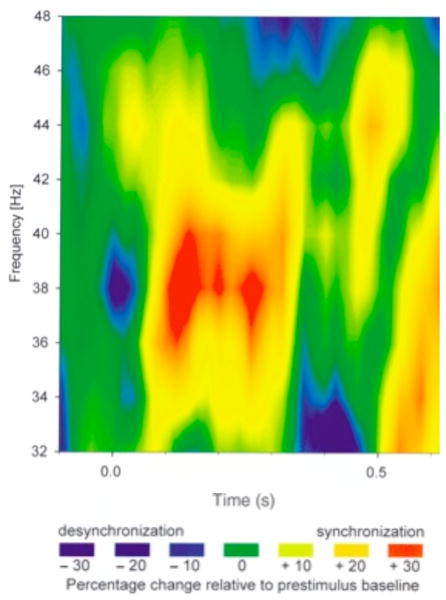

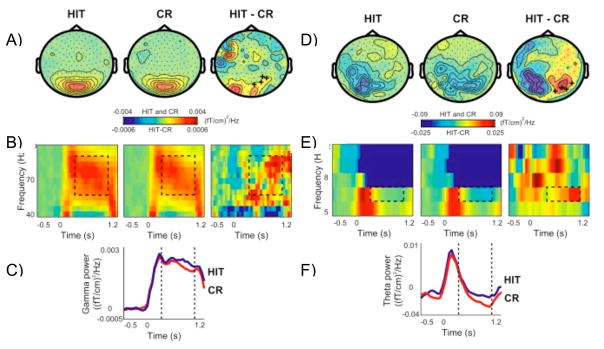
References
-
- Axmacher N, Mormann F, Fernández G, Elger CE, Fell J. Memory formation by neuronal synchronization. Brain Research Reviews. 2006;52:170–182. - PubMed
-
- Banks MI, White JA, Pearce RA. Interactions between distinct GABA(A) circuits in hippocampus. Neuron. 2000;25:449–457. - PubMed
-
- Bartos M, Vida I, Jonas P. Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nature Reviews Neuroscience. 2007;8:45–56. - PubMed
-
- Başar E, Başar-Eroglu C, Karakas S, Schürmann M. Are cognitive processes manifested in event-related gamma, alpha, theta and delta oscillations in the EEG? Neuroscience Letters. 1999;259:165–168. - PubMed
-
- Başar E, Schürmann M. Toward new theories of brain function and brain dynamics. International Journal of Psychophysiology. 2001;39:87–89. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
