

Early transformations in odor representation
- PMID: 20060600
- PMCID: PMC2839009
- DOI: 10.1016/j.tins.2009.12.004
Early transformations in odor representation
Abstract
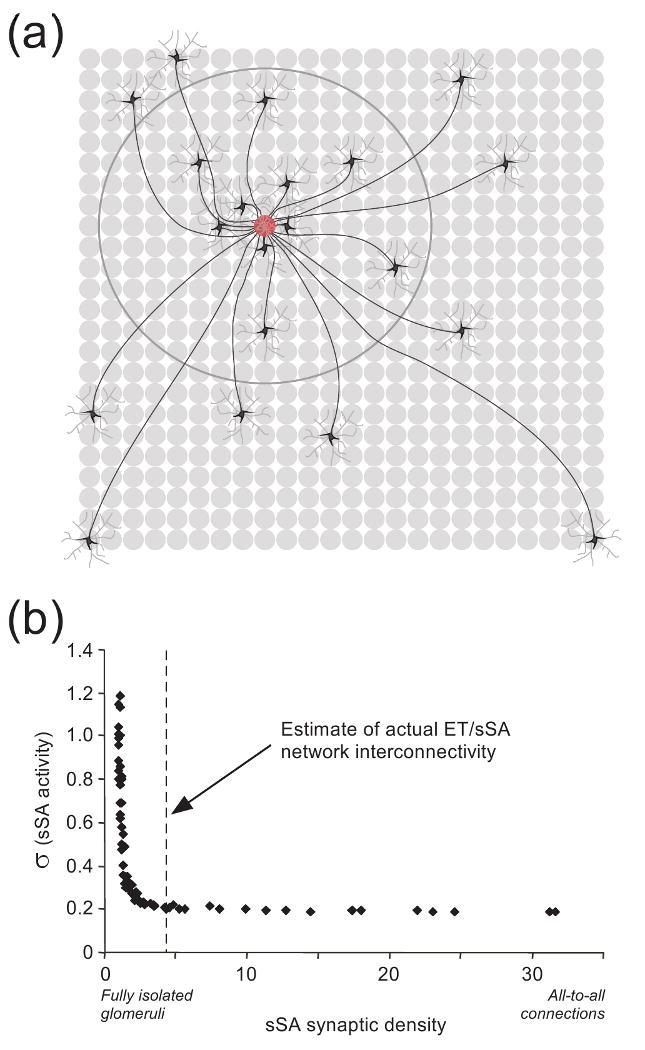
Sensory representations are repeatedly transformed by neural computations that determine which of their attributes can be effectively processed at each stage. Whereas some early computations are common across multiple sensory systems, they can utilize dissimilar underlying mechanisms depending on the properties of each modality. Recent work in the olfactory bulb has substantially clarified the neural algorithms underlying early odor processing. The high-dimensionality of odor space strictly limits the utility of topographical representations, forcing similarity-dependent computations such as decorrelation to employ unusual neural algorithms. The distinct architectures and properties of the two prominent computational layers in the olfactory bulb suggest that the bulb is directly comparable not only to the retina but also to primary visual cortex.
Copyright 2009 Elsevier Ltd. All rights reserved.
Figures


References
-
- Marr D. Vision: A computational investigation into the human representation and processing of visual information. W. H. Freeman; 1983.
-
- Marr D, Poggio T. MIT AI Memo #357. 1976. From understanding computation to understanding neural circuitry.
-
- Kleene SJ. The electrochemical basis of odor transduction in vertebrate olfactory cilia. Chem Senses. 2008;33:839–859. - PubMed
-
- Serizawa S, et al. Negative feedback regulation ensures the one receptor-one olfactory neuron rule in mouse. Science. 2003;302:2088–2094. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
