

Review
doi: 10.1016/j.sbi.2009.12.006.
Epub 2010 Jan 8.
Genome packaging in viruses
Affiliations
- PMID: 20060706
- PMCID: PMC2948483
- DOI: 10.1016/j.sbi.2009.12.006
Item in Clipboard
Review
Genome packaging in viruses
Curr Opin Struct Biol.
2010 Feb.
Abstract
Genome packaging is a fundamental process in a viral life cycle. Many viruses assemble preformed capsids into which the genomic material is subsequently packaged. These viruses use a packaging motor protein that is driven by the hydrolysis of ATP to condense the nucleic acids into a confined space. How these motor proteins package viral genomes had been poorly understood until recently, when a few X-ray crystal structures and cryo-electron microscopy (cryo-EM) structures became available. Here we discuss various aspects of genome packaging and compare the mechanisms proposed for packaging motors on the basis of structural information.
Copyright 2009 Elsevier Ltd. All rights reserved.
Figures
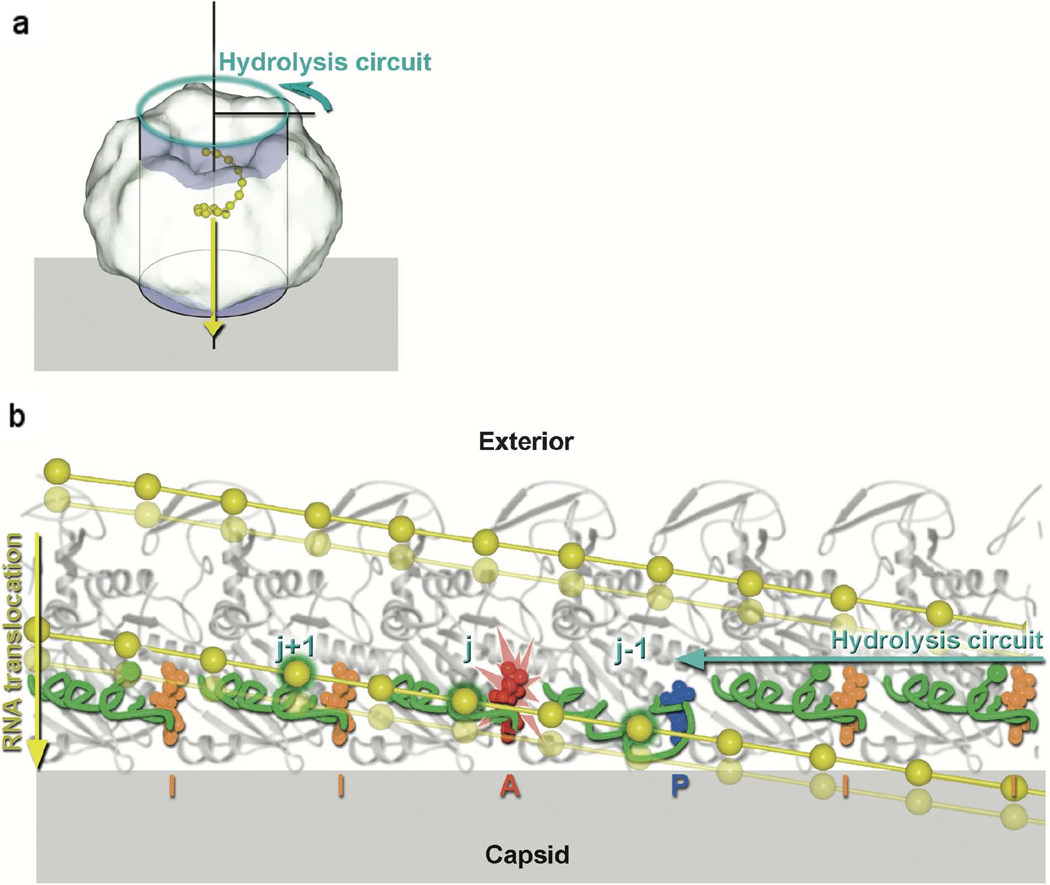
Model for translocation along P4 by the ssRNA in bacteriophage φ12 [31]. The relative movements of the RNA binding loops in six adjacent subunits as the ATP hydrolysis-induced conformational changes ripple around the ring. (a) Diagram showing the hexameric molecule represented as a cylinder associated with the viral capsid shell (gray). The direction of ssRNA (yellow) translocation is depicted by a yellow arrow, while the cyan arrow shows the direction of sequential ATP hydrolysis. To obtain the view shown in b, the coordinates are projected onto the cylinder shown and the cylinder unwrapped to lie across the page. The effect of this is that the P4 hexamer is peeled open and viewed from inside the ring looking outward. (b) Cylindrical polar projection showing a snapshot of the hexamer just prior to hydrolysis at subunit j (marked with the flash). The RNA binding loops are colored green and the ssRNA yellow with phosphates represented by balls. Phosphates interacting with the RNA binding loops are identified by a green outer glow. Diphosphate and triphosphate moieties are colored according to their conformation: AMPcPP inactive “I” (orange), AMPcPP active “A” (red), and product “P” (blue). The power stroke associated with the hydrolysis step about to occur will translocate RNA downward (from the solid RNA position to the semitransparent position). Two turns of the RNA spiral are shown. [Reprinted from Cell, Volume 118, Mancini EJ, Kainov DE, Grimes JM, Tuma R, Bamford DH, and Stuart DI: Atomic snapshots of an RNA packaging motor reveal conformational changes linking ATP hydrolysis to RNA translocation, pages 743–755, copyright 2004, with permission from Elsevier].
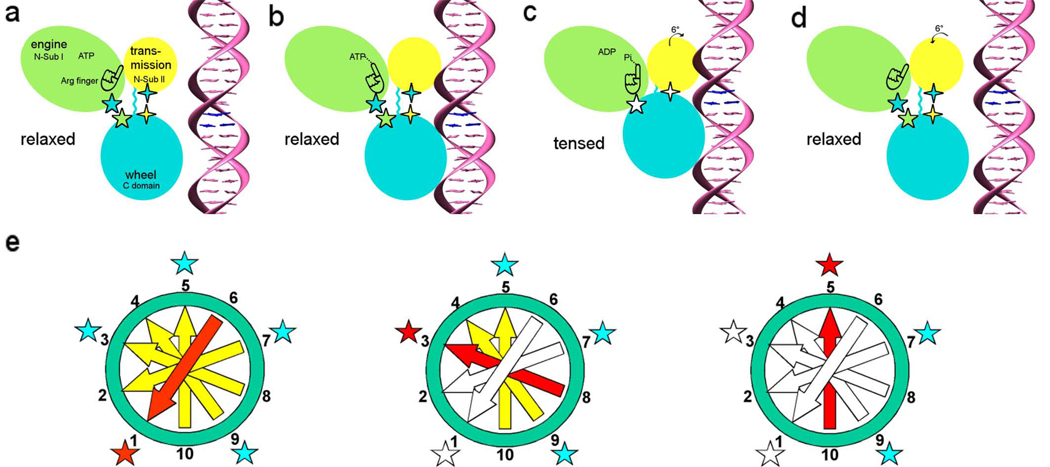
DNA packaging mechanism in bacteriophage T4 [49]. Panels (a)–(d) relate to the sequence of events that occur in a single gp17 molecule. The gp17 N-terminal subdomain I, subdomain II, and C-terminal domain are represented as green, yellow, and cyan ovals, respectively. The five-pointed stars show the charge interactions between the N-terminal subdomain I and the C-terminal domain. The four-pointed stars show the charge interaction between the N-terminal subdomain II and the C-terminal domain. The flexible linker between N- and C-terminal domain is represented by a wiggly cyan line.(a) The gp17 C-terminal domain is ready to bind DNA.(b) The C-terminal domain, when bound to the DNA, brings the DNA closer to the N-terminal domain of the same subunit. Conformational change in the N-terminal domain causes Arg162 to be placed into the ATPase active center in preparation for hydrolysis.(c) Hydrolysis of ATP has rotated the N-terminal subdomain II by about 6°, thereby aligning the charge pairs resulting in an electrostatic attraction that moves the C-terminal domain and the DNA 6.8 Å (equivalent to the distance between two base pairs) closer to the N-terminal domain and into the capsid.(d) ADP and Pi are released and the C-terminal domain returns to its original position. DNA is released and is aligned to bind the C-terminal domain of the neighboring subunit. (e) This panel relates to the synchronization of the five gp17 molecules located around the special vertex of the procapsid. Successive DNA base pairs are indicated by yellow arrows outside the procapsid, red entering the procapsid, and white inside the procapsid. The surrounding five gp17 molecules are shown as stars. The red star represents gp17 ATPase hydrolyzing ATP, the blue star represents ATPase that is ready to hydrolyze ATP, and the white star represents ATPase that has already hydrolyzed ATP. Left: Hydrolysis of ATP at position 1 translocating two base pairs into the procapsid. Middle: Hydrolysis of ATP at position 3 causing the translocation of further two base pairs into the procapsid. Right: The ATP at position 5 is ready to be hydrolyzed. [Reprinted from Cell, Volume 135, Sun S, Kondabagil K, Draper B, Alam TI, Bowman VD, Zhang Z, Hegde S, Fokine A, Rossmann MG, and Rao VB, The structure of the phage T4 DNA packaging motor suggests a mechanism dependent on electrostatic forces, pages 1251–1262, copyright 2008.]
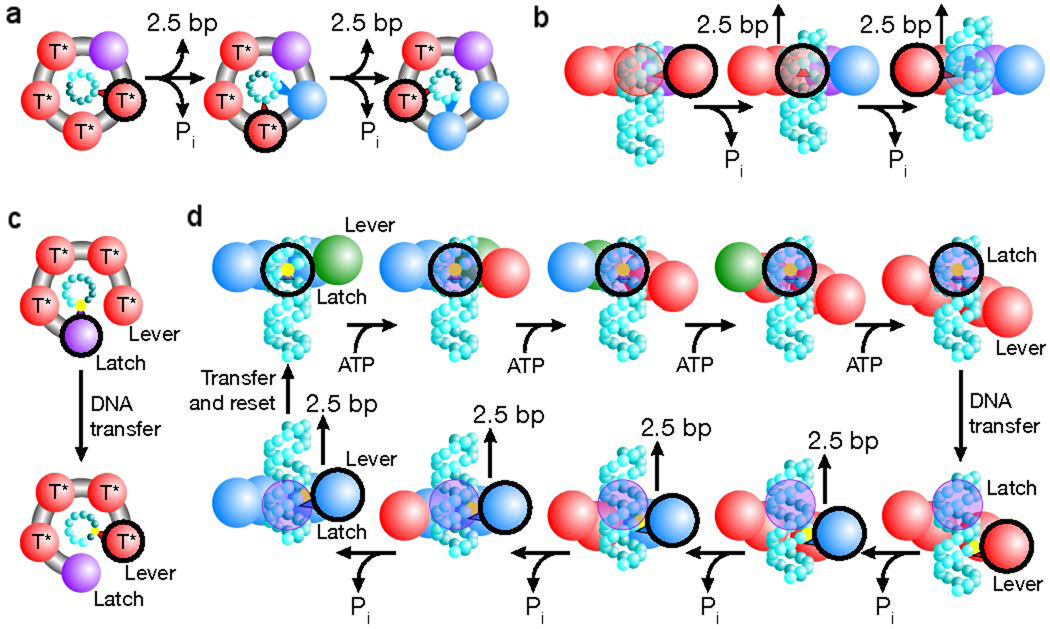
Packaging models that produce a non-integer step size in bacteriophage φ29 [60]. (a) Depiction of a translocation model in which all subunits eventually contact the DNA (cyan spheres). The contacting subunit is outlined in black (top view). (b) In such a model the size of internal conformational changes set the step size (side view). (c) Depiction of a translocation model in which only two subunits contact the DNA (black outline). (d) In such a model, one subunit maintains contact with the DNA (the latch) while the loading of each ATP introduces relative subunit–subunit rotations which distort the ring. This distortion extends one subunit (the lever) along the DNA by ~10 bp. The DNA contact point is then transferred from the latch to the lever, and the release of hydrolysis products relaxes the ring, retracting the lever and the DNA. The DNA contact is then transferred back to the latch, the ring resets and the cycle begins again. Because there are four subunits, the ring is retracted in four steps, dividing a 10 bp step into four ~2.5 bp substeps. The subunit is colored based on its substrate binding state as follows: ATP docking (green, T), tight ATP binding and activation (red, T*), ADP bound (blue) and apo (purple). [Reprinted by permission from Macmillan Publishers Ltd: Moffitt JR, Chemla YR, Aathavan K, Grimes S, Jardine PJ, Anderson DL, Bustamante C: Intersubunit coordination in a homomeric ring ATPase. Nature 2009, 457:446–450., www.nature.com ]
References
-
- Valegård K, Murray JB, Stockley PG, J SN, Liljas L. Crystal structure of an RNA bacteriophage coat protein-operator complex. Nature. 1994;371:623–626. - PubMed
-
- Fujimura T, Esteban R, Esteban LM, Wickner RB. Portable encapsidation signal of the L-A double-stranded RNA virus of S. cerevisiae. Cell. 1990;62:819–828. - PubMed
-
- Burroughs AM, Iyer LM, Aravind L. Comparative genomics and evolutionary trajectories of viral ATP dependent DNA-packaging system. Genome Dyn. 2007;3:48–65. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
