

Mechanisms of force generation by end-on kinetochore-microtubule attachments
- PMID: 20061128
- PMCID: PMC2822009
- DOI: 10.1016/j.ceb.2009.12.010
Mechanisms of force generation by end-on kinetochore-microtubule attachments
Abstract
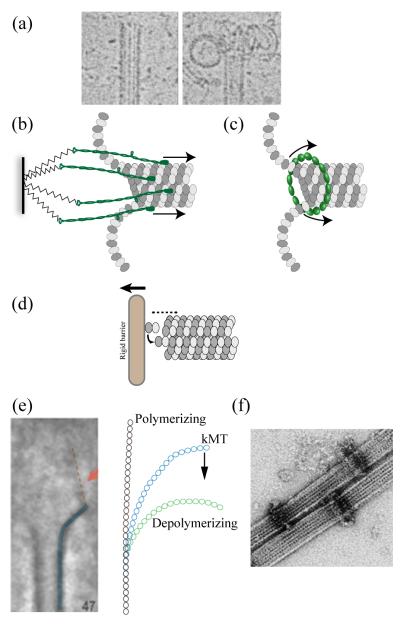
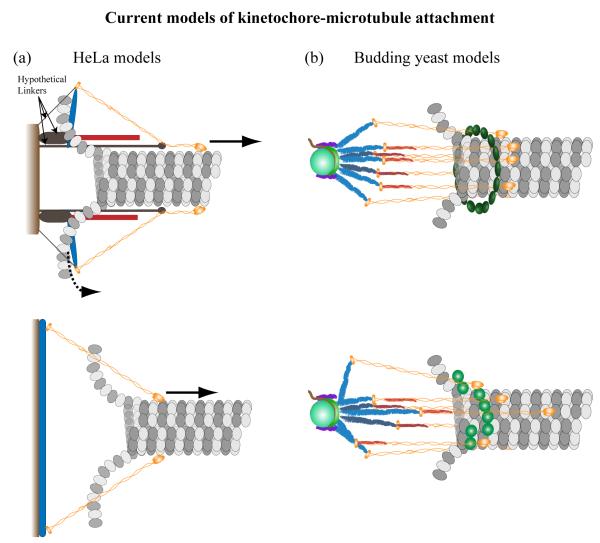
Generation of motile force is one of the main functions of the eukaryotic kinetochore during cell division. In recent years, the KMN network of proteins (Ndc80 complex, Mis12 complex, and KNL-1 complex) has emerged as a highly conserved core microtubule-binding complex at the kinetochore. It plays a major role in coupling force generation to microtubule plus-end polymerization and depolymerization. In this review, we discuss current theoretical mechanisms of force generation, and then focus on emerging information about mechanistic contributions from the Ndc80 complex in eukaryotes and the microtubule-binding Dam1/DASH complex from fungi. New information has also become available from super-resolution light microscopy on the protein architecture of the kinetochore-microtubule attachment site in both budding yeast and humans, which provides further insight into the mechanism of force generation. We briefly discuss potential contributions of motors, other microtubule-associated proteins, and microtubule depolymerases. Using the above evidence, we present speculative models of force generation at the kinetochore.
Copyright 2009 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
The conserved KMN network constitutes the core microtubule-binding site of the kinetochore.Cell. 2006 Dec 1;127(5):983-97. doi: 10.1016/j.cell.2006.09.039. Cell. 2006. PMID: 17129783
-
Stability of kinetochore-microtubule attachment and the role of different KMN network components in Drosophila.Cytoskeleton (Hoboken). 2013 Oct;70(10):661-75. doi: 10.1002/cm.21131. Epub 2013 Sep 20. Cytoskeleton (Hoboken). 2013. PMID: 23959943
-
The Hec1/Ndc80 tail domain is required for force generation at kinetochores, but is dispensable for kinetochore-microtubule attachment formation and Ska complex recruitment.Mol Biol Cell. 2020 Jul 1;31(14):1453-1473. doi: 10.1091/mbc.E20-05-0286. Epub 2020 May 13. Mol Biol Cell. 2020. PMID: 32401635 Free PMC article.
-
Kinetochore-microtubule interactions: the means to the end.Curr Opin Cell Biol. 2008 Feb;20(1):53-63. doi: 10.1016/j.ceb.2007.11.005. Epub 2008 Jan 7. Curr Opin Cell Biol. 2008. PMID: 18182282 Free PMC article. Review.
-
The KMN protein network--chief conductors of the kinetochore orchestra.J Cell Sci. 2012 Dec 15;125(Pt 24):5927-36. doi: 10.1242/jcs.093724. Epub 2013 Feb 15. J Cell Sci. 2012. PMID: 23418356 Free PMC article. Review.
Cited by
-
Ultrafast Force-Clamp Spectroscopy of Microtubule-Binding Proteins.Methods Mol Biol. 2022;2478:609-650. doi: 10.1007/978-1-0716-2229-2_22. Methods Mol Biol. 2022. PMID: 36063336 Free PMC article.
-
A second tubulin binding site on the kinesin-13 motor head domain is important during mitosis.PLoS One. 2013 Aug 28;8(8):e73075. doi: 10.1371/journal.pone.0073075. eCollection 2013. PLoS One. 2013. PMID: 24015286 Free PMC article.
-
Budding yeast kinetochore proteins, Chl4 and Ctf19, are required to maintain SPB-centromere proximity during G1 and late anaphase.PLoS One. 2014 Jul 8;9(7):e101294. doi: 10.1371/journal.pone.0101294. eCollection 2014. PLoS One. 2014. PMID: 25003500 Free PMC article.
-
Kinetochore-dependent microtubule rescue ensures their efficient and sustained interactions in early mitosis.Dev Cell. 2011 Nov 15;21(5):920-33. doi: 10.1016/j.devcel.2011.09.006. Dev Cell. 2011. PMID: 22075150 Free PMC article.
-
Deformations within moving kinetochores reveal different sites of active and passive force generation.Science. 2012 Jul 20;337(6092):355-8. doi: 10.1126/science.1221886. Epub 2012 Jun 21. Science. 2012. PMID: 22722252 Free PMC article.
References
-
-
Santaguida S, Musacchio A. The life and miracles of kinetochores. Embo J. 2009A current and comprehensive review of kinetochore protein composition, its architecture, function, and regulation. Our review is narrowly focused on the mechanisms of force generation at the kinetochore. This review provides a good discussion of the core machinery of the kinetochore.
-
-
- Cimini D. Merotelic kinetochore orientation, aneuploidy, and cancer. Biochim Biophys Acta. 2008;1786:32–40. - PubMed
-
-
Musacchio A, Salmon ED. The spindle-assembly checkpoint in space and time. Nat Rev Mol Cell Biol. 2007;8:379–393.The spindle assembly checkpoint machinery and function is reviewed in depth.
-
-
-
Hill TL. Microfilament or microtubule assembly or disassembly against a force. Proc Natl Acad Sci U S A. 1981;78:5613–5617.This paper provides an excellent discussion of the thermodynamics of force generation coupled to polymerization and depolymerization at the ends of a polymer.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
