

Cutting edge: spontaneous development of IL-17-producing gamma delta T cells in the thymus occurs via a TGF-beta 1-dependent mechanism
- PMID: 20061408
- PMCID: PMC2844788
- DOI: 10.4049/jimmunol.0903539
Cutting edge: spontaneous development of IL-17-producing gamma delta T cells in the thymus occurs via a TGF-beta 1-dependent mechanism
Abstract
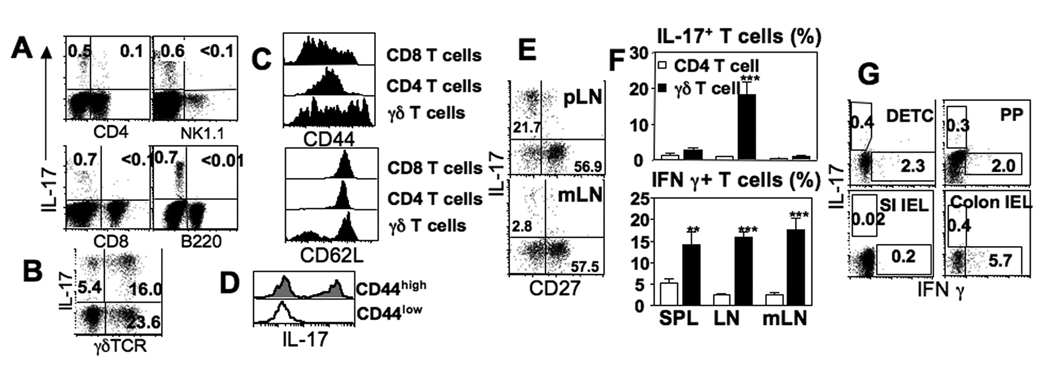
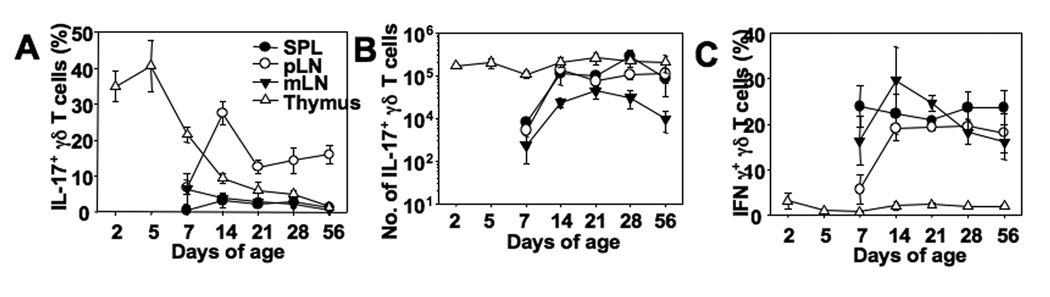
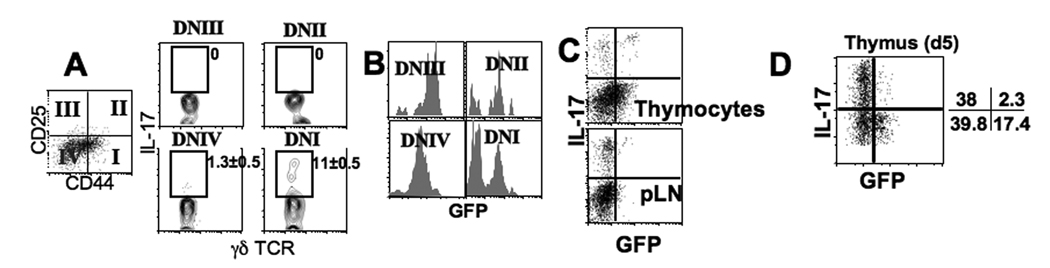
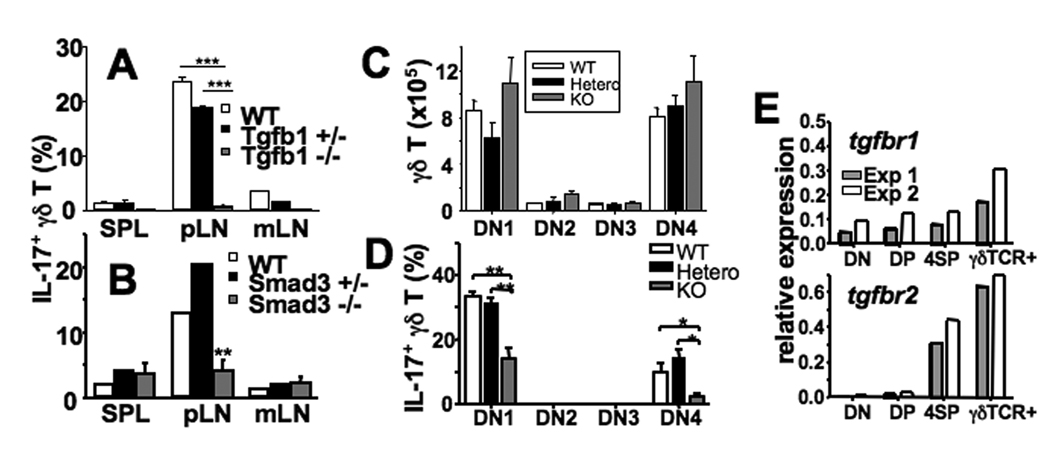
In naive animals, gammadelta T cells are innate sources of IL-17, a potent proinflammatory cytokine mediating bacterial clearance as well as autoimmunity. However, mechanisms underlying the generation of these cells in vivo remain unclear. In this study, we show that TGF-beta1 plays a key role in the generation of IL-17(+) gammadelta T cells and that it mainly occurs in the thymus particularly during the postnatal period. Interestingly, IL-17(+) gammadelta TCR(+) thymocytes were mainly CD44(high)CD25(low) cells, which seem to derive from double-negative 4 gammadelta TCR(+) cells that acquired CD44 and IL-17 expression. Our findings identify a novel developmental pathway during which IL-17-competent gammadelta T cells arise in the thymus by a TGF-beta1-dependent mechanism.
Figures
References
-
- Carding SR, Egan PJ. Gammadelta T cells: functional plasticity and heterogeneity. Nat Rev Immunol. 2002;2:336–345. - PubMed
-
- Shibata K, Yamada H, Hara H, Kishihara K, Yoshikai Y. Resident Vdelta1+ gammadelta T cells control early infiltration of neutrophils after Escherichia coli infection via IL-17 production. J Immunol. 2007;178:4466–4472. - PubMed
-
- Stark MA, Huo Y, Burcin TL, Morris MA, Olson TS, Ley K. Phagocytosis of apoptotic neutrophils regulates granulopoiesis via IL-23 and IL-17. Immunity. 2005;22:285–294. - PubMed
-
- Jensen KD, Su X, Shin S, Li L, Youssef S, Yamasaki S, Steinman L, Saito T, Locksley RM, Davis MM, Baumgarth N, Chien YH. Thymic selection determines gammadelta T cell effector fate: antigen-naive cells make interleukin-17 and antigen-experienced cells make interferon gamma. Immunity. 2008;29:90–100. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
