

The cyclic-di-GMP phosphodiesterase BinA negatively regulates cellulose-containing biofilms in Vibrio fischeri
- PMID: 20061475
- PMCID: PMC2820850
- DOI: 10.1128/JB.01048-09
The cyclic-di-GMP phosphodiesterase BinA negatively regulates cellulose-containing biofilms in Vibrio fischeri
Abstract
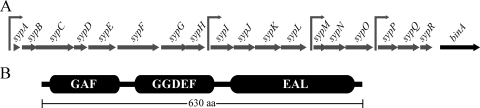
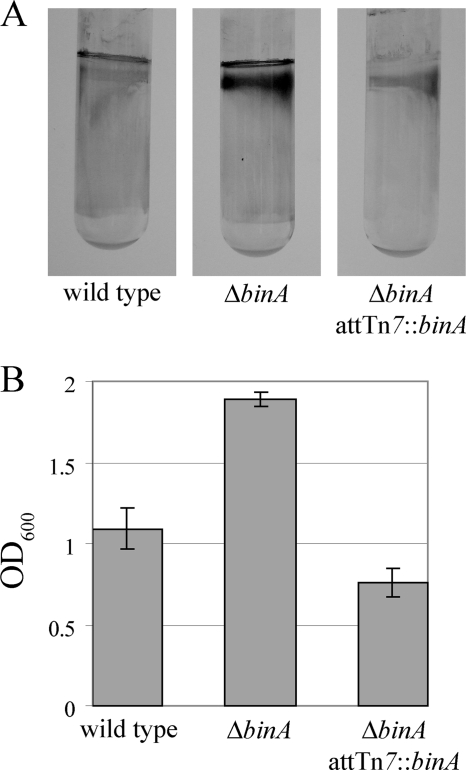
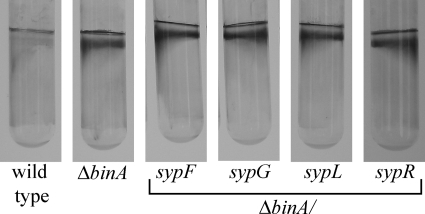
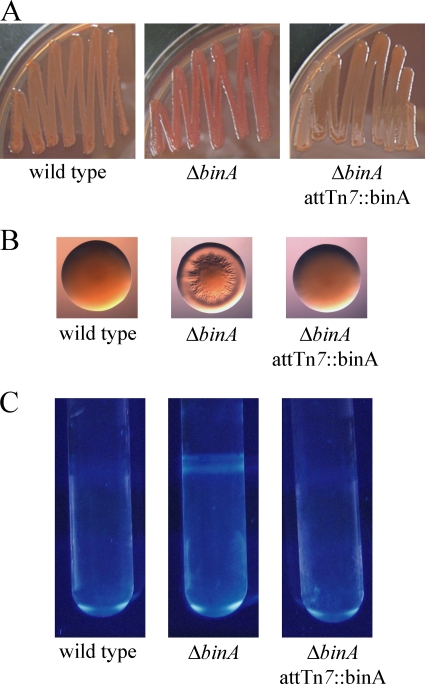
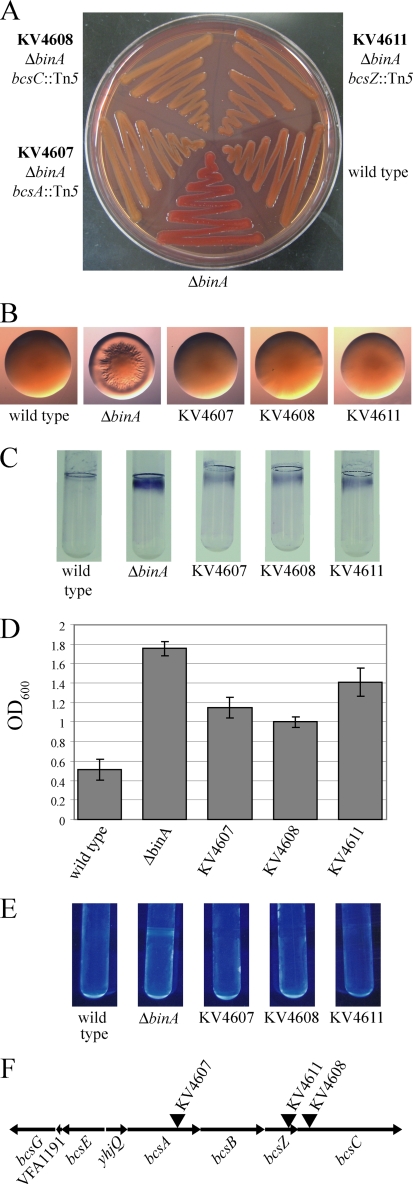
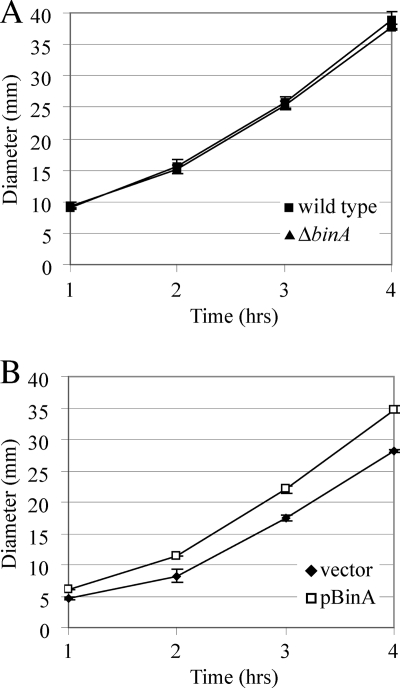
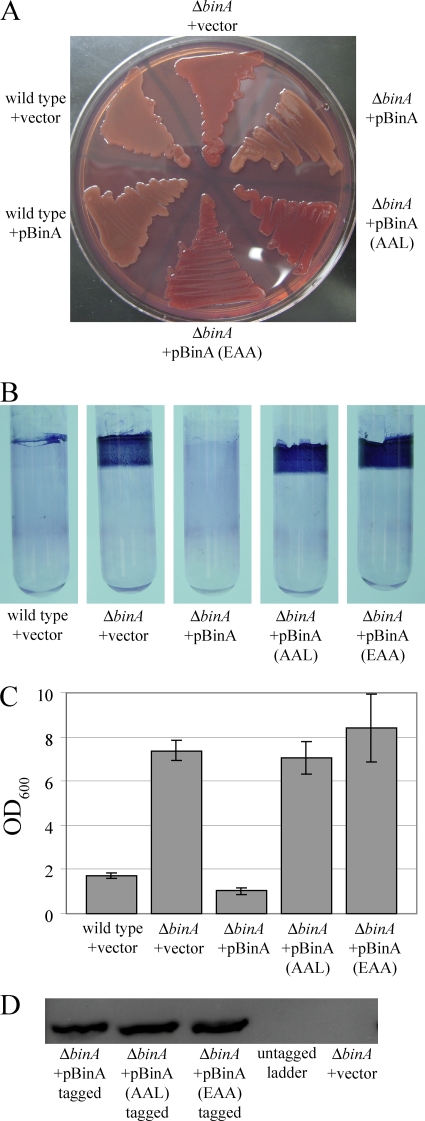
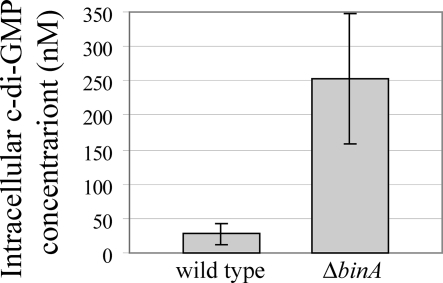
Bacteria produce different types of biofilms under distinct environmental conditions. Vibrio fischeri has the capacity to produce at least two distinct types of biofilms, one that relies on the symbiosis polysaccharide Syp and another that depends upon cellulose. A key regulator of biofilm formation in bacteria is the intracellular signaling molecule cyclic diguanylate (c-di-GMP). In this study, we focused on a predicted c-di-GMP phosphodiesterase encoded by the gene binA, located directly downstream of syp, a cluster of 18 genes critical for biofilm formation and the initiation of symbiotic colonization of the squid Euprymna scolopes. Disruption or deletion of binA increased biofilm formation in culture and led to increased binding of Congo red and calcofluor, which are indicators of cellulose production. Using random transposon mutagenesis, we determined that the phenotypes of the DeltabinA mutant strain could be disrupted by insertions in genes in the bacterial cellulose biosynthesis cluster (bcs), suggesting that cellulose production is negatively regulated by BinA. Replacement of critical amino acids within the conserved EAL residues of the EAL domain disrupted BinA activity, and deletion of binA increased c-di-GMP levels in the cell. Together, these data support the hypotheses that BinA functions as a phosphodiesterase and that c-di-GMP activates cellulose biosynthesis. Finally, overexpression of the syp regulator sypG induced binA expression. Thus, this work reveals a mechanism by which V. fischeri inhibits cellulose-dependent biofilm formation and suggests that the production of two different polysaccharides may be coordinated through the action of the cellulose inhibitor BinA.
Figures
Similar articles
-
Para-Aminobenzoic Acid, Calcium, and c-di-GMP Induce Formation of Cohesive, Syp-Polysaccharide-Dependent Biofilms in Vibrio fischeri.mBio. 2021 Oct 26;12(5):e0203421. doi: 10.1128/mBio.02034-21. Epub 2021 Oct 5. mBio. 2021. PMID: 34607467 Free PMC article.
-
High Levels of Cyclic Diguanylate Interfere with Beneficial Bacterial Colonization.mBio. 2022 Aug 30;13(4):e0167122. doi: 10.1128/mbio.01671-22. Epub 2022 Aug 2. mBio. 2022. PMID: 35916402 Free PMC article.
-
Functional analysis of cyclic diguanylate-modulating proteins in Vibrio fischeri.mSystems. 2024 Nov 19;9(11):e0095624. doi: 10.1128/msystems.00956-24. Epub 2024 Oct 22. mSystems. 2024. PMID: 39436151 Free PMC article.
-
An intricate network of regulators controls biofilm formation and colonization by Vibrio fischeri.Mol Microbiol. 2009 Nov;74(4):782-9. doi: 10.1111/j.1365-2958.2009.06899.x. Epub 2009 Oct 8. Mol Microbiol. 2009. PMID: 19818022 Free PMC article. Review.
-
Vibrio fischeri: a model for host-associated biofilm formation.J Bacteriol. 2024 Feb 22;206(2):e0037023. doi: 10.1128/jb.00370-23. Epub 2024 Jan 25. J Bacteriol. 2024. PMID: 38270381 Free PMC article. Review.
Cited by
-
Cyclic di-GMP: the first 25 years of a universal bacterial second messenger.Microbiol Mol Biol Rev. 2013 Mar;77(1):1-52. doi: 10.1128/MMBR.00043-12. Microbiol Mol Biol Rev. 2013. PMID: 23471616 Free PMC article. Review.
-
Functional analysis of cyclic diguanylate-modulating proteins in Vibrio fischeri.bioRxiv [Preprint]. 2024 Sep 24:2023.07.24.550417. doi: 10.1101/2023.07.24.550417. bioRxiv. 2024. Update in: mSystems. 2024 Nov 19;9(11):e0095624. doi: 10.1128/msystems.00956-24. PMID: 37546929 Free PMC article. Updated. Preprint.
-
A lasting symbiosis: how Vibrio fischeri finds a squid partner and persists within its natural host.Nat Rev Microbiol. 2021 Oct;19(10):654-665. doi: 10.1038/s41579-021-00557-0. Epub 2021 Jun 4. Nat Rev Microbiol. 2021. PMID: 34089008 Free PMC article. Review.
-
Arabinose induces pellicle formation by Vibrio fischeri.Appl Environ Microbiol. 2013 Mar;79(6):2069-80. doi: 10.1128/AEM.03526-12. Epub 2013 Jan 18. Appl Environ Microbiol. 2013. PMID: 23335779 Free PMC article.
-
A prototrophic suppressor of a Vibrio fischeri D-glutamate auxotroph reveals a member of the periplasmic broad-spectrum racemase family (BsrF).J Bacteriol. 2024 Mar 21;206(3):e0033323. doi: 10.1128/jb.00333-23. Epub 2024 Feb 27. J Bacteriol. 2024. PMID: 38411059 Free PMC article.
References
-
- Amikam, D., and M. Y. Galperin. 2006. PilZ domain is part of the bacterial c-di-GMP binding protein. Bioinformatics 22:3-6. - PubMed
-
- Aravind, L., and C. P. Ponting. 1997. The GAF domain: an evolutionary link between diverse phototransducing proteins. Trends Biochem. Sci. 22:458-459. - PubMed
-
- Bao, Y., D. P. Lies, H. Fu, and G. P. Roberts. 1991. An improved Tn7-based system for the single-copy insertion of cloned genes into chromosomes of Gram-negative bacteria. Gene 109:167-168. - PubMed
-
- Bobrov, A. G., O. Kirillina, and R. D. Perry. 2005. The phosphodiesterase activity of the HmsP EAL domain is required for negative regulation of biofilm formation in Yersinia pestis. FEMS Microbiol. Lett. 247:123-130. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
