

VEZF1 elements mediate protection from DNA methylation
- PMID: 20062523
- PMCID: PMC2795164
- DOI: 10.1371/journal.pgen.1000804
VEZF1 elements mediate protection from DNA methylation
Abstract
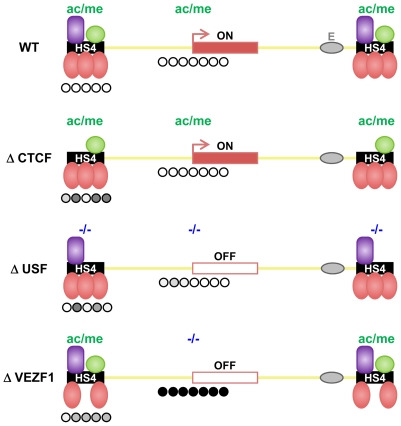
There is growing consensus that genome organization and long-range gene regulation involves partitioning of the genome into domains of distinct epigenetic chromatin states. Chromatin insulator or barrier elements are key components of these processes as they can establish boundaries between chromatin states. The ability of elements such as the paradigm beta-globin HS4 insulator to block the range of enhancers or the spread of repressive histone modifications is well established. Here we have addressed the hypothesis that a barrier element in vertebrates should be capable of defending a gene from silencing by DNA methylation. Using an established stable reporter gene system, we find that HS4 acts specifically to protect a gene promoter from de novo DNA methylation. Notably, protection from methylation can occur in the absence of histone acetylation or transcription. There is a division of labor at HS4; the sequences that mediate protection from methylation are separable from those that mediate CTCF-dependent enhancer blocking and USF-dependent histone modification recruitment. The zinc finger protein VEZF1 was purified as the factor that specifically interacts with the methylation protection elements. VEZF1 is a candidate CpG island protection factor as the G-rich sequences bound by VEZF1 are frequently found at CpG island promoters. Indeed, we show that VEZF1 elements are sufficient to mediate demethylation and protection of the APRT CpG island promoter from DNA methylation. We propose that many barrier elements in vertebrates will prevent DNA methylation in addition to blocking the propagation of repressive histone modifications, as either process is sufficient to direct the establishment of an epigenetically stable silent chromatin state.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








Similar articles
-
Histone crosstalk directed by H2B ubiquitination is required for chromatin boundary integrity.PLoS Genet. 2011 Jul;7(7):e1002175. doi: 10.1371/journal.pgen.1002175. Epub 2011 Jul 21. PLoS Genet. 2011. PMID: 21811414 Free PMC article.
-
The barrier function of an insulator couples high histone acetylation levels with specific protection of promoter DNA from methylation.Genes Dev. 2002 Jun 15;16(12):1540-54. doi: 10.1101/gad.988502. Genes Dev. 2002. PMID: 12080092 Free PMC article.
-
USF binding sequences from the HS4 insulator element impose early replication timing on a vertebrate replicator.PLoS Biol. 2012;10(3):e1001277. doi: 10.1371/journal.pbio.1001277. Epub 2012 Mar 6. PLoS Biol. 2012. PMID: 22412349 Free PMC article.
-
Chromatin domains, insulators, and the regulation of gene expression.Biochim Biophys Acta. 2012 Jul;1819(7):644-51. doi: 10.1016/j.bbagrm.2012.01.016. Epub 2012 Feb 2. Biochim Biophys Acta. 2012. PMID: 22326678 Free PMC article. Review.
-
Understanding the interplay between CpG island-associated gene promoters and H3K4 methylation.Biochim Biophys Acta Gene Regul Mech. 2020 Aug;1863(8):194567. doi: 10.1016/j.bbagrm.2020.194567. Epub 2020 Apr 29. Biochim Biophys Acta Gene Regul Mech. 2020. PMID: 32360393 Free PMC article. Review.
Cited by
-
CAGn repeat of the androgen receptor is linked to proopiomelanocortin promoter methylation-relevance for craving of male alcohol-dependent patients?Psychopharmacology (Berl). 2014 May;231(10):2059-66. doi: 10.1007/s00213-013-3349-5. Epub 2013 Nov 24. Psychopharmacology (Berl). 2014. PMID: 24271034
-
The use of chromatin insulators to improve the expression and safety of integrating gene transfer vectors.Hum Gene Ther. 2011 Jun;22(6):761-74. doi: 10.1089/hum.2010.233. Epub 2011 Mar 25. Hum Gene Ther. 2011. PMID: 21247248 Free PMC article. Review.
-
Transcription termination sequences support the expression of transgene product secreted with milk.Transgenic Res. 2019 Aug;28(3-4):401-410. doi: 10.1007/s11248-019-00122-9. Epub 2019 Mar 28. Transgenic Res. 2019. PMID: 30919251
-
Efficient Recombinase-Mediated Cassette Exchange in hPSCs to Study the Hepatocyte Lineage Reveals AAVS1 Locus-Mediated Transgene Inhibition.Stem Cell Reports. 2015 Nov 10;5(5):918-931. doi: 10.1016/j.stemcr.2015.09.004. Epub 2015 Oct 8. Stem Cell Reports. 2015. PMID: 26455413 Free PMC article.
-
The inducible tissue-specific expression of the human IL-3/GM-CSF locus is controlled by a complex array of developmentally regulated enhancers.J Immunol. 2012 Nov 1;189(9):4459-69. doi: 10.4049/jimmunol.1201915. Epub 2012 Sep 28. J Immunol. 2012. PMID: 23024272 Free PMC article.
References
-
- Kellum R, Schedl P. A position-effect assay for boundaries of higher order chromosomal domains. Cell. 1991;64:941–950. - PubMed
-
- Stalder J, Larsen A, Engel JD, Dolan M, Groudine M, et al. Tissue-specific DNA cleavages in the globin chromatin domain introduced by DNAase I. Cell. 1980;20:451–460. - PubMed
-
- Gaszner M, Felsenfeld G. Insulators: exploiting transcriptional and epigenetic mechanisms. Nat Rev Genet. 2006;7:703–713. - PubMed
-
- Valenzuela L, Kamakaka RT. Chromatin insulators. Annu Rev Genet. 2006;40:107–138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
