

Immature dengue virus: a veiled pathogen?
- PMID: 20062797
- PMCID: PMC2798752
- DOI: 10.1371/journal.ppat.1000718
Immature dengue virus: a veiled pathogen?
Abstract
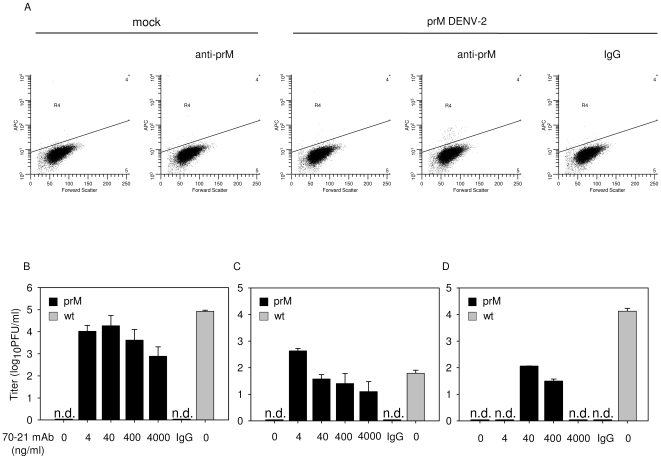
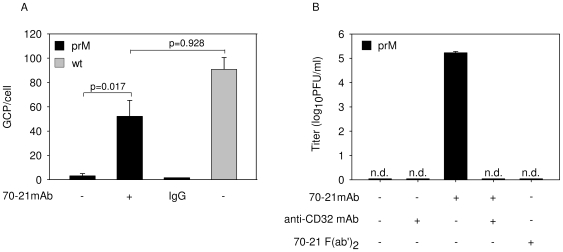
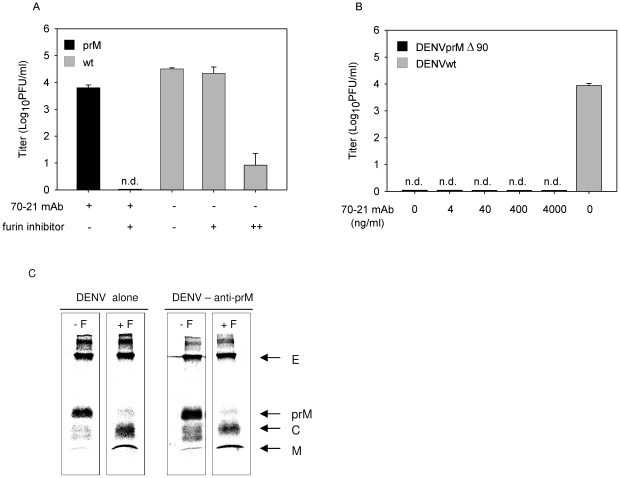
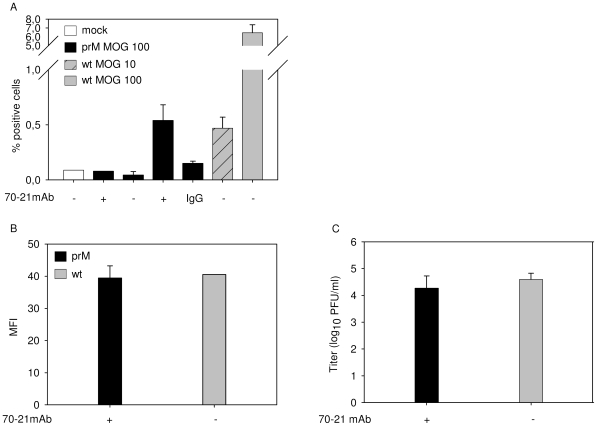
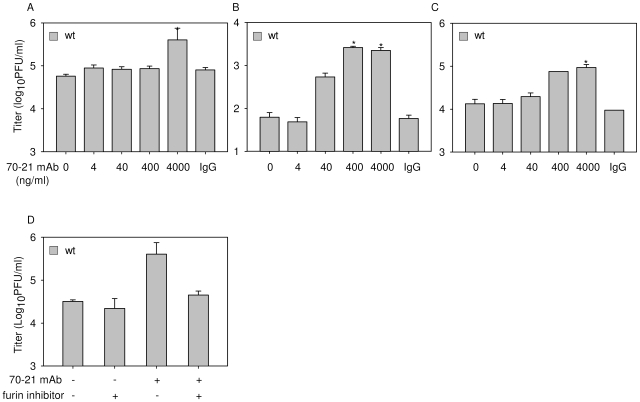
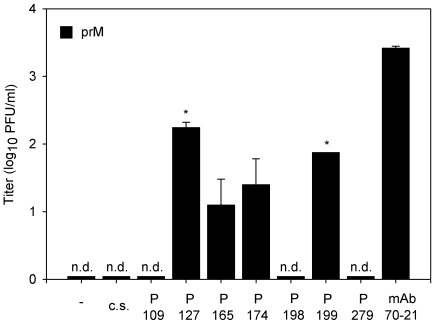
Cells infected with dengue virus release a high proportion of immature prM-containing virions. In accordance, substantial levels of prM antibodies are found in sera of infected humans. Furthermore, it has been recently described that the rates of prM antibody responses are significantly higher in patients with secondary infection compared to those with primary infection. This suggests that immature dengue virus may play a role in disease pathogenesis. Interestingly, however, numerous functional studies have revealed that immature particles lack the ability to infect cells. In this report, we show that fully immature dengue particles become highly infectious upon interaction with prM antibodies. We demonstrate that prM antibodies facilitate efficient binding and cell entry of immature particles into Fc-receptor-expressing cells. In addition, enzymatic activity of furin is critical to render the internalized immature virus infectious. Together, these data suggest that during a secondary infection or primary infection of infants born to dengue-immune mothers, immature particles have the potential to be highly infectious and hence may contribute to the development of severe disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Halstead SB. Pathogenesis of dengue: challenges to molecular biology. Science. 1988;239:476–481. - PubMed
-
- Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, et al. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J Infect Dis. 2000;181:2–9. - PubMed
-
- Kliks SC, Nimmanitya S, Nisalak A, Burke DS. Evidence that maternal dengue antibodies are important in the development of dengue hemorrhagic fever in infants. Am J Trop Med Hyg. 1988;38:411–419. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
