

Establishment of sister chromatid cohesion
- PMID: 20064425
- PMCID: PMC4867117
- DOI: 10.1016/j.cub.2009.10.067
Establishment of sister chromatid cohesion
Abstract
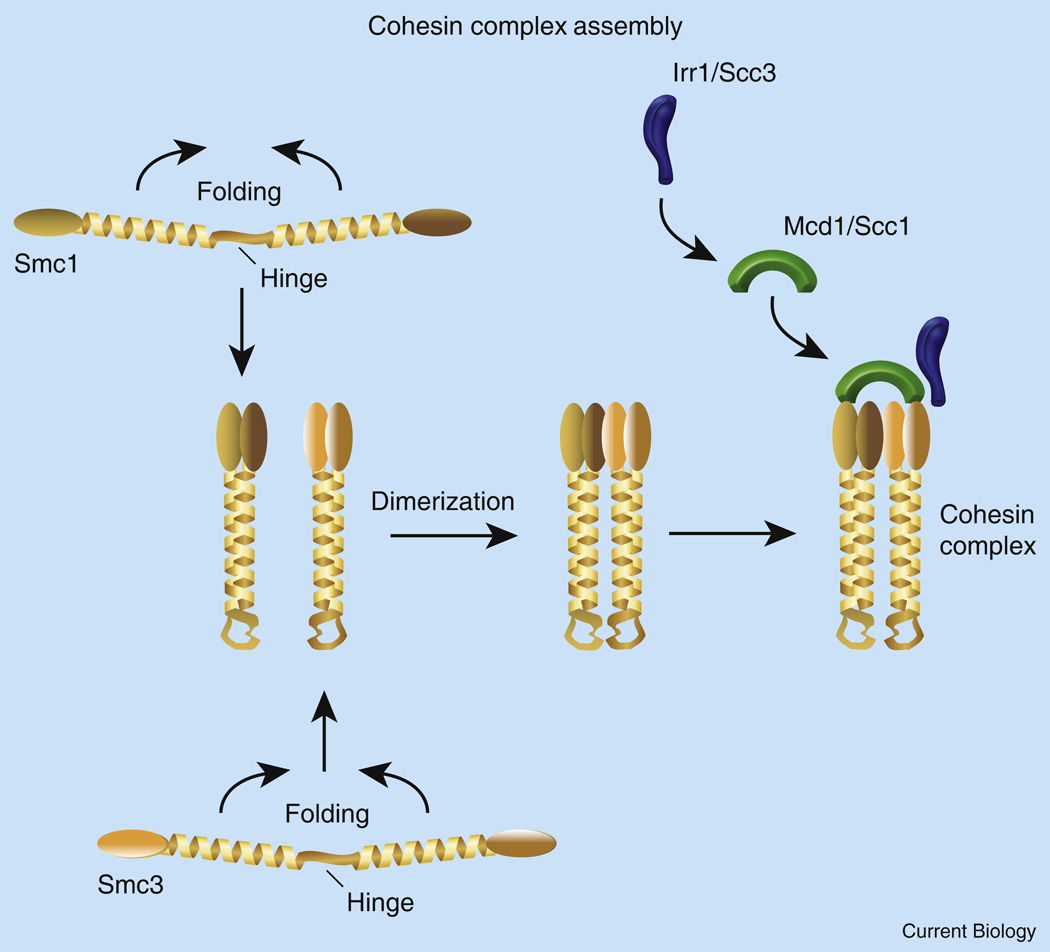
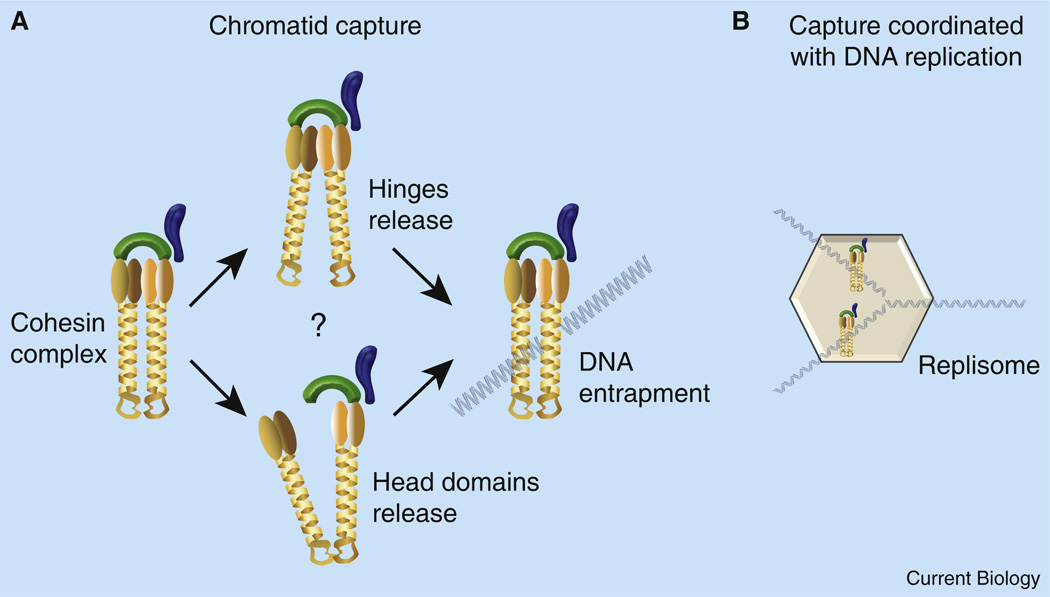
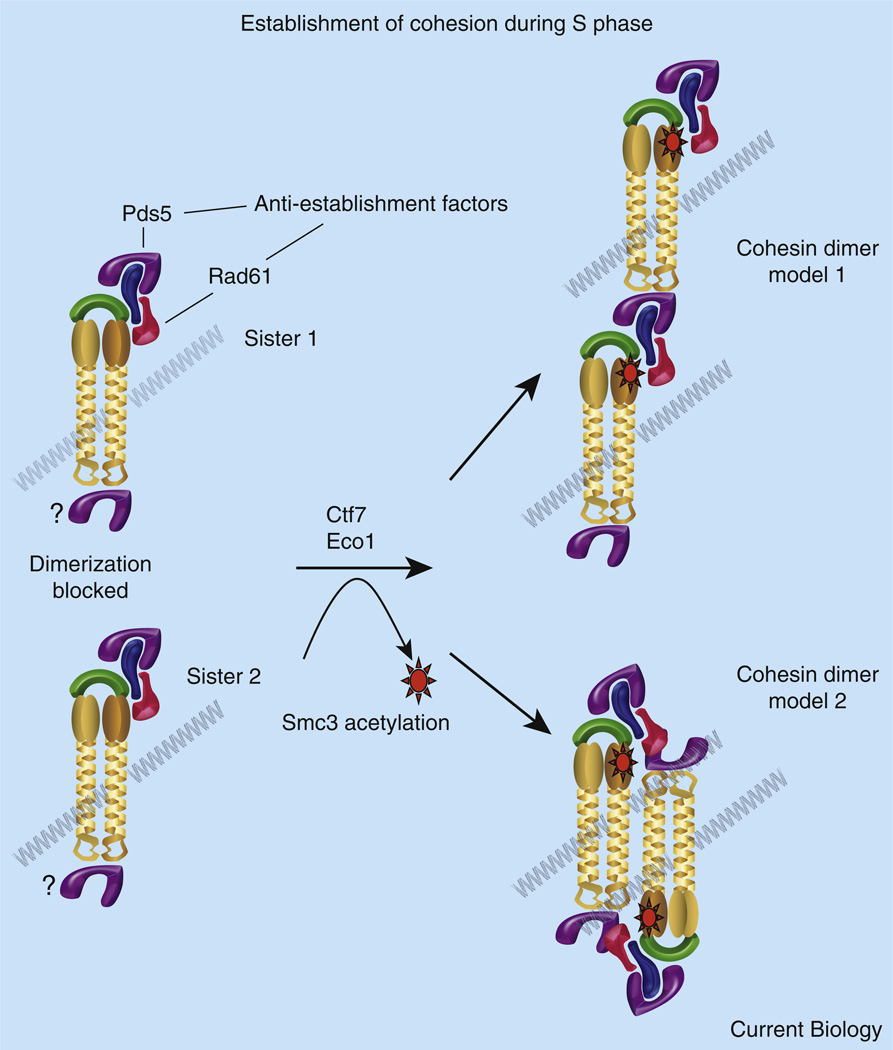
The process of sister chromatid pairing, or cohesion establishment, is coupled to DNA replication and fundamental to proper chromosome segregation and cell viability. In the past year, several articles have provided important new insights into cohesion establishment, an activity predicated on the acetyltransferase Ctf7/Eco1. Here, I review new findings that the conversion of chromatid-bound cohesins into a cohesion-competent state involves Ctf7/Eco1-mediated acetylation of the cohesin subunit Smc3. These studies further explore an anti-establishment activity that involves the binding of accessory factors WAPL/Rad61 and Pds5 to the cohesin subunit Scc3/Irr1. The anti-establishment activity of WAPL/Rad61 and Pds5 is temporarily relaxed by Ctf7/Eco1 during S phase to promote sister chromatid pairing. These findings are likely to be of clinical relevance, given the role of cohesion pathways in a wide range of disease states.
Figures
Similar articles
-
Cohesin acetylation: from antiestablishment to establishment.Mol Cell. 2009 Apr 10;34(1):1-2. doi: 10.1016/j.molcel.2009.03.011. Mol Cell. 2009. PMID: 19362531
-
The acetyltransferase Eco1 elicits cohesin dimerization during S phase.J Biol Chem. 2020 May 29;295(22):7554-7565. doi: 10.1074/jbc.RA120.013102. Epub 2020 Apr 20. J Biol Chem. 2020. PMID: 32312753 Free PMC article.
-
An Smc3 acetylation cycle is essential for establishment of sister chromatid cohesion.Mol Cell. 2010 Sep 10;39(5):689-99. doi: 10.1016/j.molcel.2010.08.008. Mol Cell. 2010. PMID: 20832721 Free PMC article.
-
Fork it over: the cohesion establishment factor Ctf7p and DNA replication.J Cell Sci. 2007 Aug 1;120(Pt 15):2471-7. doi: 10.1242/jcs.011999. J Cell Sci. 2007. PMID: 17646671 Review.
-
Mechanisms of sister chromatid pairing.Int Rev Cell Mol Biol. 2008;269:283-339. doi: 10.1016/S1937-6448(08)01005-8. Int Rev Cell Mol Biol. 2008. PMID: 18779060 Review.
Cited by
-
Reduced sister chromatid cohesion acts as a tumor penetrance modifier.PLoS Genet. 2022 Aug 22;18(8):e1010341. doi: 10.1371/journal.pgen.1010341. eCollection 2022 Aug. PLoS Genet. 2022. PMID: 35994499 Free PMC article.
-
Structural insights into the regulation of cohesion establishment by Wpl1.EMBO J. 2013 Mar 6;32(5):677-87. doi: 10.1038/emboj.2013.16. Epub 2013 Feb 8. EMBO J. 2013. PMID: 23395900 Free PMC article.
-
Inventory and phylogenetic analysis of meiotic genes in monogonont rotifers.J Hered. 2013 May-Jun;104(3):357-70. doi: 10.1093/jhered/est011. Epub 2013 Mar 13. J Hered. 2013. PMID: 23487324 Free PMC article.
-
Heterogenic final cell cycle by chicken retinal Lim1 horizontal progenitor cells leads to heteroploid cells with a remaining replicated genome.PLoS One. 2013;8(3):e59133. doi: 10.1371/journal.pone.0059133. Epub 2013 Mar 19. PLoS One. 2013. PMID: 23527113 Free PMC article.
-
Human RECQL5: guarding the crossroads of DNA replication and transcription and providing backup capability.Crit Rev Biochem Mol Biol. 2013 May-Jun;48(3):289-99. doi: 10.3109/10409238.2013.792770. Epub 2013 Apr 29. Crit Rev Biochem Mol Biol. 2013. PMID: 23627586 Free PMC article. Review.
References
-
- Skibbens RV. Mechanisms of sister chromatid pairing. Int. Rev. Cell Mol. Biol. 2009;269:283–339. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
