

CD8 T cell response and evolutionary pressure to HIV-1 cryptic epitopes derived from antisense transcription
- PMID: 20065064
- PMCID: PMC2812545
- DOI: 10.1084/jem.20092060
CD8 T cell response and evolutionary pressure to HIV-1 cryptic epitopes derived from antisense transcription
Abstract
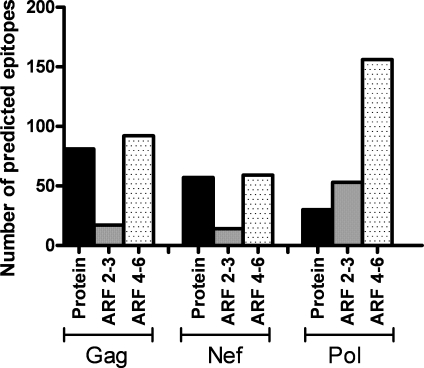
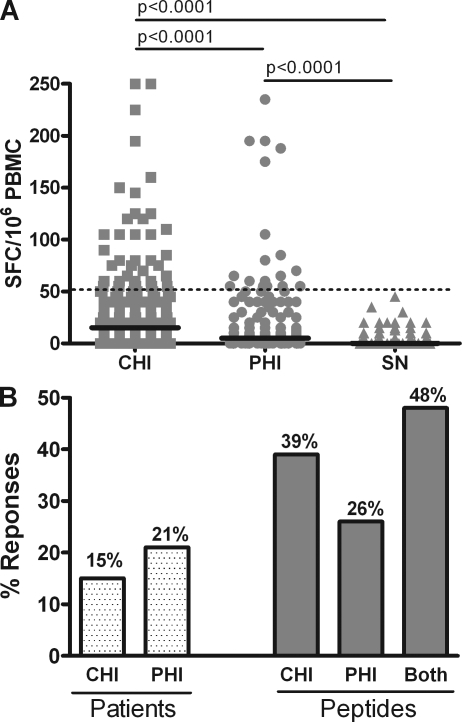
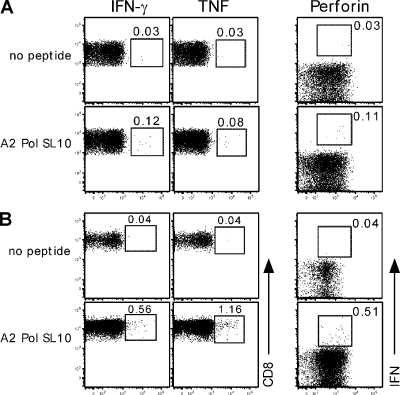
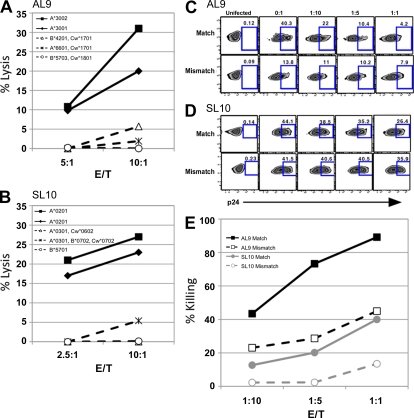
Retroviruses pack multiple genes into relatively small genomes by encoding several genes in the same genomic region with overlapping reading frames. Both sense and antisense HIV-1 transcripts contain open reading frames for known functional proteins as well as numerous alternative reading frames (ARFs). At least some ARFs have the potential to encode proteins of unknown function, and their antigenic properties can be considered as cryptic epitopes (CEs). To examine the extent of active immune response to virally encoded CEs, we analyzed human leukocyte antigen class I-associated polymorphisms in HIV-1 gag, pol, and nef genes from a large cohort of South Africans with chronic infection. In all, 391 CEs and 168 conventional epitopes were predicted, with the majority (307; 79%) of CEs derived from antisense transcripts. In further evaluation of CD8 T cell responses to a subset of the predicted CEs in patients with primary or chronic infection, both sense- and antisense-encoded CEs were immunogenic at both stages of infection. In addition, CEs often mutated during the first year of infection, which was consistent with immune selection for escape variants. These findings indicate that the HIV-1 genome might encode and deploy a large potential repertoire of unconventional epitopes to enhance vaccine-induced antiviral immunity.
Figures
References
-
- Altfeld M., Rosenberg E.S., Shankarappa R., Mukherjee J.S., Hecht F.M., Eldridge R.L., Addo M.M., Poon S.H., Phillips M.N., Robbins G.K., et al. 2001. Cellular immune responses and viral diversity in individuals treated during acute and early HIV-1 infection. J. Exp. Med. 193:169–180 10.1084/jem.193.2.169 - DOI - PMC - PubMed
-
- Bansal A., Jackson B., West K., Wang S., Lu S., Kennedy J.S., Goepfert P.A. 2008. Multifunctional T-cell characteristics induced by a polyvalent DNA prime/protein boost human immunodeficiency virus type 1 vaccine regimen given to healthy adults are dependent on the route and dose of administration. J. Virol. 82:6458–6469 10.1128/JVI.00068-08 - DOI - PMC - PubMed
-
- Betts M.R., Ambrozak D.R., Douek D.C., Bonhoeffer S., Brenchley J.M., Casazza J.P., Koup R.A., Picker L.J. 2001. Analysis of total human immunodeficiency virus (HIV)-specific CD4(+) and CD8(+) T-cell responses: relationship to viral load in untreated HIV infection. J. Virol. 75:11983–11991 10.1128/JVI.75.24.11983-11991.2001 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30 AI027767/AI/NIAID NIH HHS/United States
- P30 AI050409/AI/NIAID NIH HHS/United States
- R01 AI084772/AI/NIAID NIH HHS/United States
- P30 AI 050409/AI/NIAID NIH HHS/United States
- R21 AI073103/AI/NIAID NIH HHS/United States
- R01 AI058876/AI/NIAID NIH HHS/United States
- AI 64060/AI/NIAID NIH HHS/United States
- R01 AI071906/AI/NIAID NIH HHS/United States
- R21 AI 73103/AI/NIAID NIH HHS/United States
- R01 AI 084772/AI/NIAID NIH HHS/United States
- R01 AI064060/AI/NIAID NIH HHS/United States
- P30 AI 027767/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
