

Structural and functional roles of coevolved sites in proteins
- PMID: 20066038
- PMCID: PMC2797611
- DOI: 10.1371/journal.pone.0008591
Structural and functional roles of coevolved sites in proteins
Abstract
Background: Understanding the residue covariations between multiple positions in protein families is very crucial and can be helpful for designing protein engineering experiments. These simultaneous changes or residue coevolution allow protein to maintain its overall structural-functional integrity while enabling it to acquire specific functional modifications. Despite the significant efforts in the field there is still controversy in terms of the preferable locations of coevolved residues on different regions of protein molecules, the strength of coevolutionary signal and role of coevolution in functional diversification.
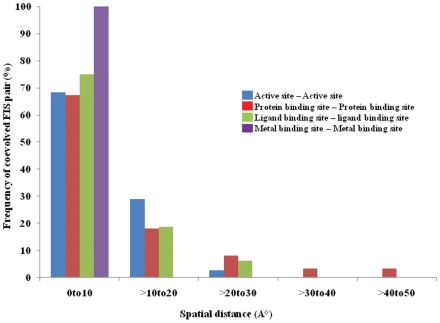
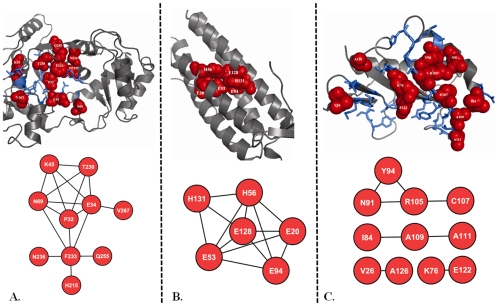
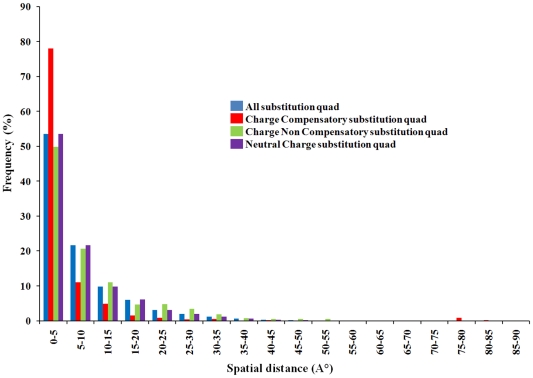
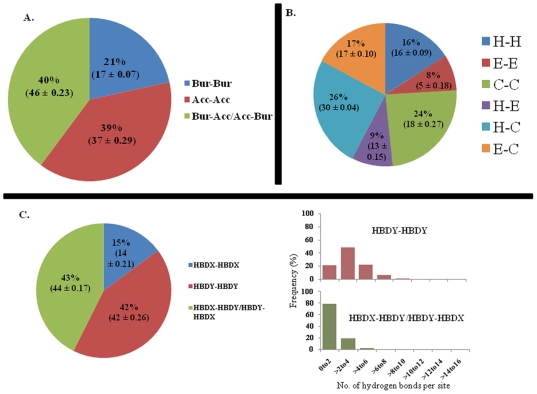
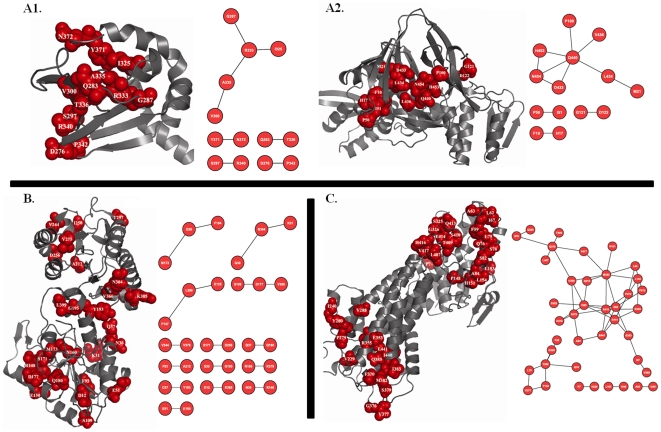
Methodology: In this paper we study the scale and nature of residue coevolution in maintaining the overall functionality and structural integrity of proteins. We employed a large scale study to investigate the structural and functional aspects of coevolved residues. We found that the networks representing the coevolutionary residue connections within our dataset are in general of 'small-world' type as they have clustering coefficient values higher than random networks and also show smaller mean shortest path lengths similar and/or lower than random and regular networks. We also found that altogether 11% of functionally important sites are coevolved with any other sites. Active sites are found more frequently to coevolve with any other sites (15%) compared to protein (11%) and ligand (9%) binding sites. Metal binding and active sites are also found to be more frequently coevolved with other metal binding and active sites, respectively. Analysis of the coupling between coevolutionary processes and the spatial distribution of coevolved sites reveals that a high fraction of coevolved sites are located close to each other. Moreover, approximately 80% of charge compensatory substitutions within coevolved sites are found at very close spatial proximity (<or= 5A), pointing to the possible preservation of salt bridges in evolution.
Conclusion: Our findings show that a noticeable fraction of functionally important sites undergo coevolution and also point towards compensatory substitutions as a probable coevolutionary mechanism within spatially proximal coevolved functional sites.
Conflict of interest statement
Figures

Similar articles
-
Coevolved residues and the functional association for intrinsically disordered proteins.Pac Symp Biocomput. 2012:140-51. Pac Symp Biocomput. 2012. PMID: 22174270
-
Coevolution in defining the functional specificity.Proteins. 2009 Apr;75(1):231-40. doi: 10.1002/prot.22239. Proteins. 2009. PMID: 18831050 Free PMC article.
-
Coevolutionary patterns in cytochrome c oxidase subunit I depend on structural and functional context.J Mol Evol. 2007 Nov;65(5):485-95. doi: 10.1007/s00239-007-9018-8. J Mol Evol. 2007. PMID: 17955155
-
Sequence-Based Protein Design: A Review of Using Statistical Models to Characterize Coevolutionary Traits for Developing Hybrid Proteins as Genetic Sensors.Int J Mol Sci. 2024 Jul 30;25(15):8320. doi: 10.3390/ijms25158320. Int J Mol Sci. 2024. PMID: 39125888 Free PMC article. Review.
-
A structural perspective of compensatory evolution.Curr Opin Struct Biol. 2014 Jun;26(100):104-12. doi: 10.1016/j.sbi.2014.05.004. Epub 2014 Jun 28. Curr Opin Struct Biol. 2014. PMID: 24981969 Free PMC article. Review.
Cited by
-
Co-evolutionary landscape at the interface and non-interface regions of protein-protein interaction complexes.Comput Struct Biotechnol J. 2021 Jun 24;19:3779-3795. doi: 10.1016/j.csbj.2021.06.039. eCollection 2021. Comput Struct Biotechnol J. 2021. PMID: 34285778 Free PMC article.
-
Duplication history and molecular evolution of the rbcS multigene family in angiosperms.J Exp Bot. 2019 Nov 18;70(21):6127-6139. doi: 10.1093/jxb/erz363. J Exp Bot. 2019. PMID: 31498865 Free PMC article.
-
Coevolution, Dynamics and Allostery Conspire in Shaping Cooperative Binding and Signal Transmission of the SARS-CoV-2 Spike Protein with Human Angiotensin-Converting Enzyme 2.Int J Mol Sci. 2020 Nov 4;21(21):8268. doi: 10.3390/ijms21218268. Int J Mol Sci. 2020. PMID: 33158276 Free PMC article.
-
Quantum coupled mutation finder: predicting functionally or structurally important sites in proteins using quantum Jensen-Shannon divergence and CUDA programming.BMC Bioinformatics. 2014 Apr 3;15:96. doi: 10.1186/1471-2105-15-96. BMC Bioinformatics. 2014. PMID: 24694117 Free PMC article.
-
Understanding the evolution of immune genes in jawed vertebrates.J Evol Biol. 2023 Jun;36(6):847-873. doi: 10.1111/jeb.14181. Epub 2023 May 31. J Evol Biol. 2023. PMID: 37255207 Free PMC article. Review.
References
-
- Kimura M. The neutral theory of molecular evolution. Cambridge: Cambridge University Press; 1983.
-
- Fitch WM. Rate of change of concomitantly variable codons. J Mol Evol. 1971;1:84–96. - PubMed
-
- Chelvanayagam G, Eggenschwiler A, Knecht L, Gonnet GH, Benner SA. An analysis of simultaneous variation in protein structures. Protein Eng. 1997;10:307–316. - PubMed
-
- Fukami-Kobayashi K, Schreiber DR, Benner SA. Detecting compensatory covariation signals in protein evolution using reconstructed ancestral sequences. J Mol Biol. 2002;319:729–743. - PubMed
-
- Choi SS, Li W, Lahn BT. Robust signals of coevolution of interacting residues in mammalian proteomes identified by phylogeny-aided structural analysis. Nat Genet. 2005;37:1367–1371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
